Мембранный потенциал нейрона. Формирование мембранного потенциала покоя
Различная концентрация веществ внутри и вне клетки является одним из важнейших признаков жизни. Всякая клетка, находящаяся в полном концентрационном равновесии с окружающей ее средой, мертва. Правда, многие вещества, в первую очередь вода, проникают в клетку по градиенту их концентраций. Однако имеется огромное количество веществ, не следующих этому правилу.
Если разделить различные вещества на классы в соответствии с их проницаемостью, то среди них можно выделить следующие основные группы.Неполярные вещества , характеризующиеся равномерным распределением электронов между атомами, образующими молекулу.Неполные полярные соединения , в которых электроны, связывающие атомы, больше притягиваются к одному из них, вследствие чего молекула превращается в диполь.Ионные соединения, или электролиты , где валентные электроны связаны лишь с одним из атомов или радикалов. В средах, обладающих высокой диэлектрической постоянной, эти связи сильно ослабляются, и отдельные части молекулы ведут себя как самостоятельные ионы. Здесь происходит диссоциация электролита на ионы: положительные - анионы и отрицательные - катионы.
Оставив в стороне вопрос о проникновении в клетку соединений двух первых классов, остановимся более подробно на характере проникновения и распределения различных ионов между клеткой и окружающей ее средой, поскольку именно это распределение лежит в основе мембранного потенциала (МП) клетки.
В настоящее время экспериментально доказано, что содержимое клетки заряжено отрицательно по отношению к ее наружной поверхности и окружающей среде. Этот заряд неодинаков у различных клеток и в большинстве случаев колеблется в пределах от -50 до -100 мв (табл. 9). Наличие электрического заряда указывает на неодинаковое распределение электролитов по обе стороны клеточной поверхностной мембраны, которое принято называть ионной асимметрией.
Таблица 9. Величины мембранных потенциалов (МП) различных возбудимых
клеток (по Латманизовой, 1965)
Каким образом возникает ионная асимметрия по разные стороны клеточной мембраны? Для выяснения этого вопроса следует хотя бы в общей форме рассмотреть поведение электролитов при прохождении через любую полупроницаемую перепонку.
Все растворенные вещества, в том числе и ионизированные, перемещаются (в случае неравномерного распределения в растворе) согласно градиенту концентрации от участков с более высокой концентрацией к участкам с меньшей концентрацией. Если участки раствора с неодинаковой концентрацией разделены какой-либо мембраной, то диффузия растворенных веществ более или менее замедляется. Ионные компоненты растворенных электролитов - анионы и катионы - нередко обладают разной проницаемостью, а следовательно, и разной подвижностью. Это связано с размерами и конфигурацией ионов, а также с величиной их гидратных оболочек. Например, образующиеся при диссоциации поваренной соли катион Na + и анион Сl - обладают различной подвижностью. Подвижность и проницаемость Na + гораздо ниже, чем Сl - . Это соответствует размерам их гидратированных диаметров 2.14 Å для Сl - и 3.4 Å для Na + . При диффузионном проникновении через мембрану из района более высокой концентрации электролита в район меньшей его концентрации ионы Сl - будут переходить на другую сторону мембраны гораздо скорее ионов Na + , так что одна из ее поверхностей на некоторое время зарядится положительно, а другая - отрицательно.
Если мембрана окажется проницаемой только для ионов Сl - и непроницаемой для ионов Na + , то возникает уже не временная, а постоянная разность потенциалов между растворами по обе стороны мембраны. Такая разность потенциалов носит названиеконцентрационной разности потенциалов и может быть рассчитана по уравнению Нернста:
![]()
где Е - разность потенциалов, в мв; R - газовая постоянная; Т - абсолютная температура; n - валентность; С 1 - более высокая, С 2 - более низкая из двух концентраций; F - число Фарадея.
Приведенные закономерности касаются того случая, когда находящиеся по обе стороны полупроницаемой мембраны растворы содержат одни и те же ионы, но в разных концентрациях.
Однако, как известно, протоплазма клетки и внеклеточной жидкости различаются не только по концентрациям, но и по составу ионов, поэтому представления о концентрационном потенциале недостаточны для объяснения событий, разыгрывающихся в районе клеточной мембраны.
Рассмотрим упрощенный случай, более близкий к реальным соотношениям электролитов в клетке. Допустим, что некая мембрана разделяет два разных электролита NaCl и КСl одинаковой концентрации:
Допустим также, что эта мембрана проницаема как для анионов Сl - , так и для обоих катионов, однако ее проницаемость для К + значительно выше, чем для Na + . Тогда К + как более подвижный ион распределяется равномерно по обе стороны мембраны гораздо скорее, чем Na + , и на все время оставания в уравнивании концентраций Na + одна из сторон мембраны, а именно та, где первоначально находился NaCl, зарядится положительно по отношению к противоположной стороне. Возникающий таким путем потенциал принято называть химическим потенциалом. Если бы мембрана была полностью непроницаемой для Na + , то этот потенциал стал бы постоянным.
В случае клеточного МП все процессы носят гораздо более сложный характер. Прежде всего электролитный состав внутриклеточной и внеклеточной среды довольно сложен и, помимо КCl и NaCl, содержит ряд других ионов, среди которых особенно важную роль в создании мембранного потенциала играют крупные органические анионы, неспособные проникать через клеточную оболочку. Приблизительные концентрации различных ионов в мышечной клетке млекопитающих приведены в табл. 10.

Таблица 10. Приблизительные концентрации ионов и потенциалы
равновесия в мышечном волокне млекопитающих и внеклеточной жидкости
(по J. W. Woodbury, 1963)
* (Рассчитано по мембранному потенциалу с помощью уравнения Нернста. )
Как видно из таблицы, внеклеточная среда характеризуется относительно высокой концентрацией ионов Na + и Сl - и низкой концентрацией ионов К + . Внутриклеточная среда, наоборот, содержит незначительные количества Na + и Сl - , но отличается высоким содержанием К + . Кроме того, внутри клетки содержится свыше 150 мМ высокомолекулярных органических анионов А - на 1 л внутриклеточной воды, по-видимому, белкового происхождения. Поэтому при построении дальнейших рассуждений необходимо принять во внимание этот дополнительный фактор.
На рис. 20 представлена очень упрощенная схема процессов, происходящих на клеточной мембране. При рассмотрении схемы делается допущение, что мембрана непроницаема не только для высокомолекулярных анионов А - , но и для ионов Na + .

Рис. 20. Формирование трансмембранного потенциала под влиянием
концентрационных градиентов. (Woodbury, 1963). Размеры символов в
правой и левой колонках указывают на относительную концентрацию
ионов во вне- и внутриклеточной жидкости. Пунктирные стрелки и
кружки - направление движения ионов, К + , A - ,
Na + и Сl - при
перемещении К + или
Сl - через
мембранные поры. Остальные объяснения в тексте
Пусть в какой-то исходный момент времени по обе стороны мембраны среда с растворенными в ней ионами электрически нейтральна, т. е. положительные и отрицательные частицы по обе стороны мембраны уравновешивают друг друга. Проследим поведение частиц, способных проникать через "мембранные поры". * Ввиду высокой внутриклеточной концентрации ионов К + эти ионы будут диффундировать через клеточную мембрану изнутри кнаружи в соответствии с градиентом концентрации. Уравновешивающие электрический заряд ионов К + высокомолекулярные анионы не могут их сопровождать ввиду того, что мембрана для них непроницаема. Ионы Na + также не могут заменить их во внутриклеточной среде, так как, согласно допущению, они не проникают через мембрану снаружи внутрь. В результате возникает частичное разделение зарядов вокруг мембраны. Наружная ее сторона в результате появившегося избытка калия во внеклеточной среде начинает заряжаться положительно, а внутренняя сторона из-за появления не нейтрализованных калием высокомолекулярных ионов А - заряжается отрицательно. В районе мембраны возникает электрическое поле, которое начинает вмешиваться в происходящий процесс, поскольку диффундирующие ионы К + имеют заряд. Как известно, одноименные электрические заряды отталкиваются, поэтому возникающий во внеклеточной среде положительный заряд начинает противодействовать дальнейшему поступлению ионов К + изнутри клетки наружу. Таким образом, диффузия К + из клетки наружу вызывает возникновение электростатических сил, препятствующих первичному диффузионному процессу, т. е. создается отрицательная обратная связь, ограничивающая выход К + из клетки. Когда положительный потенциал внешней среды по отношению к соответственно нарастающему внутриклеточному отрицательному потенциалу достигает определенной величины, устанавливается динамическое равновесие между числом выходящим из клетки ионов К + и числом этих ионов, входящих в клетку. Соответствующую этому моменту разность потенциалов по обе стороны мембраны обычно обозначают какпотенциал равновесия для данного иона . Из хода рассуждений ясно, что его величина зависит от соотношений внутренней и наружной концентрации этого иона и может быть вычислена из уравнения Нернста.
* (Выражение "мембранные поры" не следует понимать буквально, как отверстия определенной величины. Неодинаковая проницаемость мембраны к различным частицам, по-видимому, связана со сложным взаимодействием между молекулами мембраны и этими частицами. Тем не менее размер проникающих частиц, как будет видно из дальнейшего изложения, играет существенную роль. )
Аналогичные рассуждения можно провести в отношении иона Сl - . Этот анион начинает поступать в условный исходный момент снаружи внутрь в связи с его относительно высокой концентрацией во внеклеточной жидкости. При этом внутри клетки создается отрицательный заряд, а на наружной поверхности мембраны - положительный. Как и в случае с К + , создается отрицательная обратная связь, в конце концов ограничивающая поступление ионов Сl - внутрь клетки по достижении потенциала равновесия между наружной и внутренней средой.
Естественно, что процесс выхода К + из клетки и процесс вхождения в клетку ионов Сl - находится во взаимодействии и уровень устанавливающейся разности потенциалов определяется совокупностью происходящих событий.
На приведенной схеме не трудно разобраться в некоторых механизмах формирования МП покоящейся живой клетки. * Количественные расчеты с использованием уравнения Нернста показывают, что ионы Сl - и К + распределяются по обе стороны мембраны приблизительно в соответствии с величиной МП, так что электрические и концентрационные градиенты уравновешивают друг друга.
* (Покоящейся условно называется клетка, не реализующая своей специфической деятельности. Как будет видно из дальнейшего, понятие покоя является физиологической абстракцией. )
Действительно описанный механизм возникновения МП мог бы иметь место, если бы мембрана клетки была полностью непроницаема для ионов Na + . Однако изящные и строгие исследования Ходжкина и Кейнса (Keynes, 1954; Hodgkin a. Keynes, 1955) с использованием радиоактивного изотопа натрия Na24 показали, что клеточная мембрана проницаема для Na + , хотя его проницаемость приблизительно в 50 раз ниже проницаемости для К + и Сl - . На икроножной мышце лягушки экспериментально было показано (Keynes, 1954), что за секунду через 1 см 2 клеточной поверхности внутрь мышечного волокна проникает около 10 -11 M Na + . При таком "втекании" Na + внутрь клетки без соответствующего "вытекания" из нее его внутриклеточная концентрация в течение часа должна была бы удвоиться. Однако этого на самом деле не происходит, хотя и концентрационный и электрический градиенты "понуждают" Na + поступать внутрь клетки. Следовательно, должны существовать какие-то специальные механизмы, обеспечивающие прохождение ионов Na + через мембрану против электрохимического градиента, т. е. изнутри наружу. Эти механизмы получили название активного переноса. Для осуществления последнего необходима постоянная затрата энергии, вырабатывающейся в результате клеточных метаболических процессов.
К сожалению, до сих пор остается невыясненным, каким образом энергия клеточного метаболизма используется при активном переносе, однако имеются достаточно четкие экспериментальные данные об участии в этом процессе аденозинтрифосфата и некоторых других фосфатов (Caldwell a. Keynes, 1957; Caldwell, Hodgkin, Keynes a. Shaw, 1960a; Hogdkin, 1964).
Прямую связь между активным переносом и обменом веществ впервые показали Ходжкин и Кейнс (Hodgkin a. Keynes, 1955) в своих исследованиях на гигантском аксоне каракатицы (sepia). Методика исследования основывалась на использовании радиоизотопа натрия - Na 24 и позволила выявить ряд интересных закономерностей активного переноса. Прежде всего оказалось, что ингибиторы обмена веществ (динитрофенол, цианид, азид), блокирующие определенные звенья обменных окислительных реакций, почти полностью подавляют процесс выведения Na + из клетки * . Аналогично действует снижение температуры препарата, резко уменьшающее уровень обменных процессов (рис. 21).
* (В последнее время показано также блокирующее действие азида на активный перенос ионов Na + в мотонейронах спинного мозга кошек (Ito a. Oshima, 1964). )

Рис. 21. Выход ионов Na + из
гигантского аксона (каракатицы) при изменении состава омывающего
раствора добавлением динитрофенола (ДНФ) (А) и изменении температуры
(Б) (Hodgkin a. Keynes, 1955). По оси абсцисс - время, в мин.; по
оси ординат - выход радиоактивного Na 24 из
клетки, измеренный в импульсах в мин. Общий выход N + (радиоактивного
и обычного) пропорционален выходу радиоактивного Na + ,
исключая те изменения, которые возникают в результате постепенного
разведения радиоактивного Na + обычным
ввиду естественных ионнообменных процессов (это постепенное падение
концентрации радиоактивного Na + показано
в графе "морская вода")
Далее было обнаружено, что выведение Na + в общем пропорционально его внутренней концентрации и не зависит от его наружной концентрации. В то же время выход Na + резко уменьшается при снижении концентрации калия во внешней среде и увеличивается при повышении внешней концентрации К + . Эта зависимость также показана на рис. 21. Кроме того, выяснилось, что метаболические яды снижают также поступление К + из внешней среды внутрь аксона и что этот процесс блокируется указанными воздействиями почти в такой же степени, как выведение Na + из клетки. Все это позволило сделать заключение, что механизм активного переноса работает по принципу электронейтральности, "вынося" ионы Na + из клетки и "внося" в клетку эквивалентное количество ионов К + (Hodgkin, 1958). *
* (Позднее было обнаружено, что связь между выведением Na + и введением внутрь клетки К + при активном переносе довольно свободная, во всяком случае не абсолютно жесткого типа (один к одному), как это предполагалось вначале (Caldwell, Hodgkin, Keynes a. Shaw, 1960a, 1960b). До последнего времени считалось, что активный перенос определяется уровнем внутренней концентрации Na + (Hodkin a. Keynes, 1956; Hodgkin, 1958). В последних работах Экклса и его сотрудников приводятся данные о том, что калиевый компонент активного переноса активируется снижением внутренней концентрации К и что при этом внутрь клетки активно переносится КСl (Eccles, 1964; Eccles, Eccles a. Ito, 1964). )
Возникает вопрос, как может механизм, основанный на принципе электронейтральности, создавать поляризацию мембраны. Это не трудно понять, если учесть, что внутри клетки имеет место значительное количество высокомолекулярных анионов А - и что проницаемость ионов Na + в 50 раз ниже проницаемости ионов К + . Будучи выведен из клетки механизмом активного переноса, натрий благодаря диффузии возвращается в клетку в 50 раз медленнее, чем введенный внутрь клетки калий выходит из нее в виде диффузионного потока. К тому же калий диффундирует из клетки в сопровождении нейтрализующего его иона Сl - . В результате перечисленных процессов абсолютное количество положительных ионов внутри клетки убывает: активно выводится Na + и пассивно - сопровождаемый ионами Сl - калий. В клетке возникает избыток отрицательных зарядов благодаря наличию высокомолекулярных ионов А - .
Это, конечно, весьма упрощенная система рассуждений. Тем не менее она в общих чертах отражает существо явлений, не касаясь некоторых усложняющих деталей, например, вопроса о распределении в этих условиях воды.
Существует много различных гипотез, в которых делается попытка объяснить механизм активного переноса, однако ни одна из них не доказана экспериментально. Наиболее привлекательны схемы с так называемым "промежуточным носителем". В качестве примера можно привести схему Шоу-Глинна (Glynn, 1957) (рис. 22) Преимущество такой схемы заключается в том, что она не предполагает жесткой связи между выведением Na + и поступлением в клетку К + . Так, сохранение частичного выведения Na + при полном отсутствии К + во внешней среде может быть объяснено тем, что Na + имеет также некоторое сродство и к субстанциих . Гипотетическая схема должна объяснять такую возможность, поскольку выведение Na + действительно частично сохраняется даже при полном отсутствии К + во внеклеточной среде.

Рис. 22. Гипотетическая схема калий-натриевого обменного "насоса". (Glynn,
1957). Предполагается, что субстанции X и Y функционируют внутри
мембраны. X обладает большим сродством к К + ; Y обладает
большим сродством к Na + . X и У перемещаются через
мембрану только в комбинации с соответствующим ионом
До сих пор все явления и механизмы, связанные с генерацией МП, в целях простоты изложения описывались лишь с качественной стороны. Однако для более ясного представления о происходящих процессах целесообразно привести некоторые количественные характеристики поляризации мембраны, тем более что это позволит также составить более четкое представление об объеме происходящих ионообменных процессов. *
* (Нижеследующий цифровой материал взят из "Нейрофизиологии" Т. Ру и соавторов (Ruch et al., 1963). )
Емкость мембраны поперечнополосатого мышечного волокна лягушки определена в эксперименте и составляет около 10 мкф на см 2 . Величина заряда мембраны - этого биологического конденсатора - определяется величиной разности потенциалов по обе стороны мембраны (Е м) и ее емкостью (С м): q = C м ⋅ Е м. Е м примем за 90 мв. Тогда заряд q = 10 × 10 -6 ф/см 2 × 0.09 в = 9 × 10 -7 кулонов на 1 см 2 . Если пересчитать полученную величину заряда на число ионов, участвующих в его формировании, то получаются следующие результаты.
По закону Авогадро 1 граммолекула содержит 6.023 × 10 23 молекул. В данном случае мы имеем дело с одновалентными ионами, несущими на себе 1 ед. заряда, равную по величине заряду одного электрона. Заряд электрона, выраженный в кулонах, составляет 1.6 × 10 -19 . Тогда заряд одного моля одновалентных катионов составляет 1.6 × 10 -19 × 6.023 × 10 23 = 96500 кулонов. Поскольку заряд на клеточной мембране мышечного волокна составляет 9 × 10 -7 кулонов см 2 , то это означает, что эта мембрана на участке 1 см 2 разделяет всего 9 × 10 -7 /96500 = 9.5 × 10 -12 моля ионизированных одновалентных частиц. В то же время в 1 см 3 внутриклеточной жидкости содержится 1.5 × 10 -6 моля катионов или анионов, т. е. в несколько миллионов раз больше. Подсчеты показывают, что слой внутриклеточной жидкости толщиной всего в 6 Å обеспечивает достаточное количество ионов, чтобы создать на мембране потенциал в 90 мв.
Эти величины поясняют, какое незначительное количество ионов от их общего числа в клетке принимает участие в поддержании МП и, как это будет показано ниже, в генерации потенциала действия.
Следует также отметить, что клетка располагает достаточными энергетическими возможностями для осуществления активного переноса Na + . Кейнс и Майзель (Keynes a. Maisel, 1954) экспериментально показали, что расход энергии на активный перенос Na + в скелетной мышце лягушки значительно меньше общих энергетических расходов клетки, рассчитанных по потреблению кислорода. * В большинстве опытов в покоящейся мышце только около 10% энергетических расходов идет на активное выведение Na + . Если принять, что коэффициент полезного действия при активном переносе составляет только 50% или даже меньше, то все равно энергетический расход на этот процесс образует относительно небольшую часть энергии, вырабатываемой клеткой. Эти количественные данные существенно подкрепляют концепцию механизма активного переноса, показывая, что с точки зрения энергетического баланса клетки подобный процесс вполне возможен.
17 ..Статья на конкурс «био/мол/текст»: Потенциал покоя - это важное явление в жизни всех клеток организма, и важно знать, как он формируется. Однако это сложный динамический процесс, трудный для восприятия целиком, особенно для студентов младших курсов (биологических, медицинских и психологических специальностей) и неподготовленных читателей. Впрочем, при рассмотрении по пунктам, вполне возможно понять его основные детали и этапы. В работе вводится понятие потенциала покоя и выделяются основные этапы его формирования с использованием образных метафор, помогающих понять и запомнить молекулярные механизмы формирования потенциала покоя.
Мембранные транспортные структуры - натрий-калиевые насосы - создают предпосылки для возникновения потенциала покоя. Предпосылки эти - разность в концентрации ионов на внутренней и наружной сторонах клеточной мембраны. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка ионов калия (K +) выровнять свою концентрацию по обе стороны мембраны приводит к его утечке из клетки и потере вместе с ними положительных электрических зарядов, за счёт чего значительно усиливается общий отрицательный заряд внутренней поверхности клетки. Эта «калиевая» отрицательность составляет бóльшую часть потенциала покоя (−60 мВ в среднем), а меньшую его часть (−10 мВ) составляет «обменная» отрицательность, вызванная электрогенностью самого ионного насоса-обменника.
Давайте разбираться подробнее.
Зачем нам нужно знать, что такое потенциал покоя и как он возникает?
Вы знаете, что такое «животное электричество»? Откуда в организме берутся «биотоки»? Как живая клетка, находящаяся в водной среде, может превратиться в «электрическую батарейку» и почему она моментально не разряжается?
На эти вопросы можно ответить только в том случае, если узнать, как клетка создаёт себе разность электрических потенциалов (потенциал покоя) на мембране.
Совершенно очевидно, что для понимания того, как работает нервная система, необходимо вначале разобраться, как работает её отдельная нервная клетка - нейрон. Главное, что лежит в основе работы нейрона - это перемещение электрических зарядов через его мембрану и появление вследствие этого на мембране электрических потенциалов. Можно сказать, что нейрон, готовясь к своей нервной работе, вначале запасает энергию в электрической форме, а затем использует ее в процессе проведения и передачи нервного возбуждения.
Таким образом, наш самый первый шаг к изучению работы нервной системы - это понять, каким образом появляется электрический потенциал на мембране нервных клеток. Этим мы и займёмся, и назовём этот процесс формированием потенциала покоя .
Определение понятия «потенциал покоя»
В норме, когда нервная клетка находится в физиологическом покое и готова к работе, у неё уже произошло перераспределение электрических зарядов между внутренней и наружной сторонами мембраны. За счёт этого возникло электрическое поле, и на мембране появился электрический потенциал - мембранный потенциал покоя .
Таким образом, мембрана оказывается поляризованной. Это означает, что она имеет разный электрический потенциал наружной и внутренней поверхностей. Разность между этими потенциалами вполне возможно зарегистрировать.
В этом можно убедиться, если ввести внутрь клетки микроэлектрод, соединённый с регистрирующей установкой. Как только электрод попадает внутрь клетки, он мгновенно приобретает некоторый постоянный электроотрицательный потенциал по отношению к электроду, расположенному в окружающей клетку жидкости. Величина внутриклеточного электрического потенциала у нервных клеток и волокон, например, гигантских нервных волокон кальмара, в покое составляет около −70 мВ. Эту величину называют мембранным потенциалом покоя (МПП). Во всех точках аксоплазмы этот потенциал практически одинаков.
Ноздрачёв А.Д. и др. Начала физиологии .
Ещё немного физики. Макроскопические физические тела, как правило, электрически нейтральны, т.е. в них в равных количествах содержатся как положительные, так и отрицательные заряды. Зарядить тело можно, создав в нем избыток заряженных частиц одного вида, например, трением о другое тело, в котором при этом образуется избыток зарядов противоположного вида. Учитывая наличие элементарного заряда (e ), полный электрический заряд любого тела можно представить как q = ±N×e , где N - целое число.
Потенциал покоя - это разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны, когда клетка находится в состоянии физиологического покоя. Его величина измеряется изнутри клетки, она отрицательна и составляет в среднем −70 мВ (милливольт), хотя в разных клетках может быть различной: от −35 мВ до −90 мВ.
Важно учитывать, что в нервной системе электрические заряды представлены не электронами, как в обычных металлических проводах, а ионами - химическими частицами, имеющими электрический заряд. И вообще в водных растворах в виде электрического тока перемещаются не электроны, а ионы. Поэтому все электрические токи в клетках и окружающей их среде - это ионные токи .
Итак, изнутри клетка в покое заряжена отрицательно, а снаружи - положительно. Это свойственно всем живым клеткам, за исключением, разве что, эритроцитов, которые, наоборот, заряжены отрицательно снаружи. Если говорить конкретнее, то получается, что снаружи вокруг клетки будут преобладать положительные ионы (катионы Na + и K +), а внутри - отрицательные ионы (анионы органических кислот, не способные свободно перемещаться через мембрану, как Na + и K +).
Теперь нам всего лишь осталось объяснить, каким же образом всё получилось именно так. Хотя, конечно, неприятно сознавать, что все наши клетки кроме эритроцитов только снаружи выглядят положительными, а внутри они - отрицательные.
Термин «отрицательность», который мы будем применять для характеристики электрического потенциала внутри клетки, пригодится нам для простоты объяснения изменений уровня потенциала покоя. В этом термине ценно то, что интуитивно понятно следующее: чем больше отрицательность внутри клетки - тем ниже в отрицательную сторону от нуля смещён потенциал, а чем меньше отрицательность - тем ближе отрицательный потенциал к нулю. Это намного проще понять, чем каждый раз разбираться в том, что же именно означает выражение «потенциал возрастает» - возрастание по абсолютному значению (или «по модулю») будет означать смещение потенциала покоя вниз от нуля, а просто «возрастание» - смещение потенциала вверх к нулю. Термин «отрицательность» не создаёт подобных проблем неоднозначности понимания.
Сущность формирования потенциала покоя
Попробуем разобраться, откуда берётся электрический заряд нервных клеток, хотя их никто не трёт, как это делают физики в своих опытах с электрическими зарядами.
Здесь исследователя и студента поджидает одна из логических ловушек: внутренняя отрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а, наоборот, из-за потери некоторого количества положительных частиц (катионов)!
Так куда же деваются из клетки положительно заряженные частицы? Напомню, что это покинувшие клетку и скопившиеся снаружи ионы натрия - Na + - и калия - K + .
Главный секрет появления отрицательности внутри клетки
Сразу откроем этот секрет и скажем, что клетка лишается части своих положительных частиц и заряжается отрицательно за счёт двух процессов:
- вначале она обменивает «свой» натрий на «чужой» калий (да-да, одни положительные ионы на другие, такие же положительные);
- потом из неё происходит утечка этих «наменянных» положительных ионов калия, вместе с которыми из клетки утекают положительные заряды.
Эти два процесса нам и надо объяснить.
Первый этап создания внутренней отрицательности: обмен Na + на K +
В мембране нервной клетки постоянно работают белковые насосы-обменники (аденозинтрифосфатазы, или Na + /K + -АТФазы), встроенные в мембрану. Они меняют «собственный» натрий клетки на наружный «чужой» калий.
Но ведь при обмене одного положительного заряда (Na +) на другой такой же положительный заряд (K +) никакого дефицита положительных зарядов в клетке возникать не может! Правильно. Но, тем не менее, из-за этого обмена в клетке остаётся очень мало ионов натрия, потому что они почти все ушли наружу. И в то же время клетка переполняется ионами калия, которые в неё накачали молекулярные насосы. Если бы мы могли попробовать на вкус цитоплазму клетки, мы бы заметили, что в результате работы насосов-обменников она превратилась из солёной в горько-солёно-кислую, потому что солёный вкус хлорида натрия сменился сложным вкусом довольно-таки концентрированного раствора хлорида калия. В клетке концентрация калия достигает 0,4 моль/л. Растворы хлорида калия в пределах 0,009–0,02 моль/л имеют сладкий вкус, 0,03–0,04 - горький, 0,05–0,1 - горько-солёный, а начиная с 0,2 и выше - сложный вкус, состоящий из солёного, горького и кислого .
Важно здесь то, что обмен натрия на калий - неравный . За каждые отданные клеткой три иона натрия она получает всего два иона калия . Это приводит к потере одного положительного заряда при каждом акте ионного обмена. Так что уже на этом этапе за счёт неравноценного обмена клетка теряет больше «плюсов», чем получает взамен. В электрическом выражении это составляет примерно −10 мВ отрицательности внутри клетки. (Но помните, что нам надо ещё найти объяснение для оставшихся −60 мВ!)
Чтобы легче было запомнить работу насосов-обменников, образно можно выразиться так: «Клетка любит калий!» Поэтому клетка и затаскивает калий к себе, несмотря на то, что его и так в ней полно. И поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия. И поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. (Вот что делает любовь, пусть она даже и не настоящая!)
Кстати, интересно, что клетка не рождается с готовым потенциалом покоя. Ей его ещё надо создать. Например, при дифференцировке и слиянии миобластов потенциал их мембраны изменяется от −10 до −70 мВ, т.е. их мембрана становится более отрицательной - поляризуется в процессе дифференцировки. А в экспериментах на мультипотентных мезенхимальных стромальных клетках костного мозга человека искусственная деполяризация, противодействующая потенциалу покоя и уменьшающая отрицательность клеток, даже ингибировала (угнетала) дифференцировку клеток .
Образно говоря, можно выразиться так: Создавая потенциал покоя, клетка «заряжается любовью». Это любовь к двум вещам:
- любовь клетки к калию (поэтому клетка насильно затаскивает его к себе);
- любовь калия к свободе (поэтому калий покидает захватившую его клетку).
Механизм насыщения клетки калием мы уже объяснили (это работа насосов-обменников), а механизм ухода калия из клетки объясним ниже, когда перейдём к описанию второго этапа создания внутриклеточной отрицательности. Итак, результат деятельности мембранных ионных насосов-обменников на первом этапе формирования потенциала покоя таков:
- Дефицит натрия (Na +) в клетке.
- Избыток калия (K +) в клетке.
- Появление на мембране слабого электрического потенциала (−10 мВ).
Можно сказать так: на первом этапе ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Второй этап создания отрицательности: утечка ионов K + из клетки
Итак, что начинается в клетке после того, как с ионами поработают её мембранные натрий-калиевые насосы-обменники?
Из-за образовавшегося дефицита натрия внутри клетки этот ион при каждом удобном случае норовит устремиться внутрь : растворённые вещества всегда стремятся выровнять свою концентрацию во всём объёме раствора. Но это у натрия получается плохо, поскольку ионные натриевые каналы обычно закрыты и открываются только при определённых условиях: под воздействием специальных веществ (трансмиттеров) или при уменьшении отрицательности в клетке (деполяризации мембраны).
В то же время в клетке имеется избыток ионов калия по сравнению с наружной средой - потому что насосы мембраны насильно накачали его в клетку. И он, тоже стремясь уравнять свою концентрацию внутри и снаружи, норовит, напротив, выйти из клетки . И это у него получается!
Ионы калия K + покидают клетку под действием химического градиента их концентрации по разные стороны мембраны (мембрана значительно более проницаема для K + , чем для Na +) и уносят с собой положительные заряды. Из-за этого внутри клетки нарастает отрицательность.
Тут ещё важно понять то, что ионы натрия и калия как бы «не замечают» друг друга, они реагируют только «на самих себя». Т.е. натрий реагирует на концентрацию натрия же, но «не обращает внимания» на то, сколько вокруг калия. И наоборот, калий реагирует только на концентрацию калия и «не замечает» натрий. Получается, что для понимания поведения ионов надо по отдельности рассматривать концентрации ионов натрия и калия. Т.е. надо отдельно сравнить концентрацию по натрию внутри и снаружи клетки и отдельно - концентрацию по калию внутри и снаружи клетки, но не имеет смысла сравнивать натрий с калием, как это, бывает, делается в учебниках.
По закону выравнивания химических концентраций, который действует в растворах, натрий «хочет» снаружи войти в клетку; туда же его влечёт и электрическая сила (как мы помним, цитоплазма заряжена отрицательно). Хотеть-то он хочет, но не может, так как мембрана в обычном состоянии плохо его пропускает. Натриевые ионные каналы, имеющиеся в мембране, в норме закрыты. Если все же его заходит немножко, то клетка сразу же обменивает его на наружный калий с помощью своих натрий-калиевых насосов-обменников. Получается, что ионы натрия проходят через клетку как бы транзитом и не задерживаются в ней. Поэтому натрий в нейронах всегда в дефиците.
А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Он выходит наружу через особые каналы в мембране - «калиевые каналы утечки», которые в норме открыты и выпускают калий .
К + -каналы утечки постоянно открыты при нормальных значениях мембранного потенциала покоя и проявляют взрывы активности при сдвигах мембранного потенциала, которые длятся несколько минут и наблюдаются при всех значениях потенциала. Усиление К + -токов утечки ведёт к гиперполяризации мембраны, тогда как их подавление - к деполяризации. ...Однако, существование канального механизма, ответственного за токи утечки, долгое время оставалось под вопросом. Только сейчас стало ясно, что калиевая утечка - это ток через специальные калиевые каналы.
Зефиров А.Л. и Ситдикова Г.Ф. Ионные каналы возбудимой клетки (структура, функция, патология) .
От химического - к электрическому
А теперь - ещё раз самое главное. Мы должны осознанно перейти от движения химических частиц к движению электрических зарядов .
Калий (K +) положительно заряжен, и поэтому он, когда выходит из клетки, выносит из неё не только самого себя, но и положительный заряд. За ним изнутри клетки к мембране тянутся «минусы» - отрицательные заряды. Но они не могут просочиться через мембрану - в отличие от ионов калия - т.к. для них нет подходящих ионных каналов, и мембрана их не пропускает. Помните про оставшиеся необъяснёнными нами −60 мВ отрицательности? Это и есть та самая часть мембранного потенциала покоя, которую создаёт утечка ионов калия из клетки! И это - большая часть потенциала покоя.
Для этой составной части потенциала покоя есть даже специальное название - концентрационный потенциал . Концентрационный потенциал - это часть потенциала покоя, созданная дефицитом положительных зарядов внутри клетки, образовавшимся за счёт утечки из неё положительных ионов калия .
Ну, а теперь немного физики, химии и математики для любителей точности.
Электрические силы связаны с химическими по уравнению Гольдмана. Его частным случаем является более простое уравнение Нернста , по формуле которого можно рассчитать трансмембранную диффузионную разность потенциалов на основе различной концентрации ионов одного вида по разные стороны мембраны. Так, зная концентрацию ионов калия снаружи и внутри клетки, можно рассчитать калиевый равновесный потенциал E K:

где Е к - равновесный потенциал, R - газовая постоянная, Т - абсолютная температура, F - постоянная Фарадея, К + внеш и K + внутр - концентрации ионов К + снаружи и внутри клетки, соответственно. По формуле видно, что для расчёта потенциала между собой сравниваются концентрации ионов одного вида - K + .
Более точно итоговая величина суммарного диффузионного потенциала, который создаётся утечкой нескольких видов ионов, рассчитывается по формуле Гольдмана-Ходжкина-Катца. В ней учтено, что потенциал покоя зависит от трех факторов: (1) полярности электрического заряда каждого иона; (2) проницаемости мембраны Р для каждого иона; (3) [концентраций соответствующих ионов] внутри (внутр) и снаружи мембраны (внеш). Для мембраны аксона кальмара в покое отношение проводимостей Р K: PNa :P Cl = 1: 0,04: 0,45 .
Заключение
Итак, поте нциал покоя состоит из двух частей:
- −10 мВ , которые получаются от «несимметричной» работы мембранного насоса-обменника (ведь он больше выкачивает из клетки положительных зарядов (Na +), чем закачивает обратно с калием).
- Вторая часть - это всё время утекающий из клетки калий, уносящий положительные заряды. Его вклад - основной: −60 мВ . В сумме это и дает искомые −70 мВ.
Что интересно, калий перестанет выходить из клетки (точнее, его вход и выход уравниваются) только при уровне отрицательности клетки −90 мВ. В этом случае сравняются химические и электрические силы, проталкивающие калий через мембрану, но направляющие его в противоположные стороны. Но этому мешает постоянно подтекающий в клетку натрий, который несёт с собой положительные заряды и уменьшает отрицательность, за которую «борется» калий. И в итоге в клетке поддерживается равновесное состояние на уровне −70 мВ.
Вот теперь мембранный потенциал покоя окончательно сформирован.
Схема работы Na + /K + -АТФазы наглядно иллюстрирует «несимметричный» обмен Na + на K + : выкачивание избыточного «плюса» в каждом цикле работы фермента приводит к отрицательному заряжению внутренней поверхности мембраны. Чего в этом ролике не сказано, так это того, что АТФаза ответственна за менее чем 20% потенциала покоя (−10 мВ): оставшаяся «отрицательность» (−60 мВ) появляется за счет выхода из клетки через «калиевые каналы утечки» ионов K + , стремящихся выровнять свою концентрацию внутри клетки и вне нее.
Литература
- Jacqueline Fischer-Lougheed, Jian-Hui Liu, Estelle Espinos, David Mordasini, Charles R. Bader, et. al.. (2001). Human Myoblast Fusion Requires Expression of Functional Inward Rectifier Kir2.1 Channels . J Cell Biol . 153 , 677-686;
- Liu J.H., Bijlenga P., Fischer-Lougheed J. et al. (1998). Role of an inward rectifier K + current and of hyperpolarization in human myoblast fusion . J. Physiol. 510 , 467–476;
- Sarah Sundelacruz, Michael Levin, David L. Kaplan. (2008). Membrane Potential Controls Adipogenic and Osteogenic Differentiation of Mesenchymal Stem Cells . PLoS ONE . 3 , e3737;
- Павловская М.В. и Мамыкин А.И. Электростатика. Диэлектрики и проводники в электрическом поле. Постоянный ток / Электронное пособие по общему курсу физики. СПб: Санкт-Петербургский государственный электротехнический университет;
- Ноздрачёв А.Д., Баженов Ю.И., Баранникова И.А., Батуев А.С. и др. Начала физиологии: Учебник для вузов / Под ред. акад. А.Д. Ноздрачёва. СПб: Лань, 2001. - 1088 с.;
- Макаров А.М. и Лунева Л.А. Основы электромагнетизма / Физика в техническом университете. Т. 3;
- Зефиров А.Л. и Ситдикова Г.Ф. Ионные каналы возбудимой клетки (структура, функция, патология). Казань: Арт-кафе, 2010. - 271 с.;
- Родина Т.Г. Сенсорный анализ продовольственных товаров. Учебник для студентов вузов. М.: Академия, 2004. - 208 с.;
- Кольман Я. и Рем К.-Г. Наглядная биохимия. М.: Мир, 2004. - 469 с.;
- Шульговский В.В. Основы нейрофизиологии: Учебное пособие для студентов вузов. М.: Аспект Пресс, 2000. - 277 с..
Нервная система
регулирует деятельность
организма благодаря
проведению информации
(возбуждения) по сети
нервных клеток.
Цель нейрофизиологии –
это понять биологические
механизмы, которые лежат
в основе проведения
информации по нервной
системе.
Нейроны проводят информацию на большие
расстояния с помощью электрических сигналов,
которые распространяются по аксону.
Используется специальный тип электрических
сигналов – нервный импульс или потенциал
действия.
Потенциал действия является основным
носителем информации в нервной системе
Мембранный потенциал покоя нейрона
Процесс генерации и распространения ПДпроисходит на мембране нейрона.
Клетки, которые способны генерировать и проводить
нервный импульс, имеют возбудимую мембрану.
Мембранный потенциал покоя нейрона
Если на нейрон не действует раздражитель, то оннаходится в состоянии покоя.
В состоянии покоя внешняя сторона мембраны
нейрона заряжена положительно, а внутренняя –
отрицательно. Это состояние называется
мембранным потенциалом покоя.
Мембранный потенциал покоя (МПП) – это
разность потенциалов на мембране нейрона, которую
нейрон имеет в состоянии относительного
физиологического покоя.
Мембранный потенциал покоя нейрона
Потенциал действия – это кратковременноеизменение мембранного потенциала, при котором
внешняя сторона мембраны на одну тысячную
секунду становится отрицательной, а внутренняя –
положительной.
Мембранный потенциал покоя нейрона
1.2.
3.
Чтобы понять, как нейрон передает информацию,
необходимо изучить:
каким образом в состоянии покоя на мембране
нейрона возникает и поддерживается мембранный
потенциал покоя;
каким образом мембранный потенциал
кратковременно изменяется во время генерации
нервного импульса;
каким образом нервный импульс распространяется
вдоль мембраны нейрона.
Мембранный потенциал покоя нейрона
Механизм возникновения МППДвижение ионов
МПП возникает в результате движения ионов
(заряженных частиц) через ионные каналы
мембраны клетки.
Ионы – это атомы или молекулы, которые имеют
положительный (катионы) или отрицательный
(анионы) заряд.
Например, K+, Na+, Cl¯, Ca2+ и т.д.
Механизм возникновения МПП
Движение ионов черезионные каналы связано с
действием двух факторов:
1. диффузия
2. электрическая сила
Диффузия – это движение
ионов из мест c высокой
концентрацией в места с
низкой концентрацией.
Механизм возникновения МПП
ТерминыГрадиент концентрации – это разность
концентрации ионов.
Сила концентрационного градиента – это сила
химической природы, которая перемещает ионы из
мест с высокой концентрацией в места с низкой
концентрацией данного иона.
Правило: чем больше градиент концентрации, тем
больше сила концентрационного градиента.
10. Механизм возникновения МПП
Электрическая сила (I) – этосила, которая перемещает
ионы в электрическом поле.
Электрическая сила
перемещает отрицательные
ионы (анионы) к
положительному заряду
(аноду), а положительные ионы
(катионы) – к отрицательному
заряду (катоду).
11. Механизм возникновения МПП
Движение электрических зарядов в электрическомполе называется электрическим током.
Сила электрического тока определяется двумя
факторами:
1. электрическим потенциалом
2. электрической проводимостью
12. Механизм возникновения МПП
Электрический потенциал (V) – этосила, которая отражает различия в
заряде между катодом и анодом.
Чем больше различия в заряде, тем
больше электрический потенциал, тем
сильнее ток ионов.
Электрический потенциал измеряется
в Вольтах (V).
Электрическая проводимость – это
относительная способность
электрических зарядов двигаться в
электрическом поле.
Чем выше электрическая
проводимость, тем сильнее ток ионов.
13. Механизм возникновения МПП
Электрическое сопротивление (R) – сила,препятствующая движению электрических зарядов.
Электрическое сопротивление измеряется в Омах
(Ω) .
Соотношение между электрическим потенциалом,
сопротивлением и силой тока описывается законом
Ома.
I = V/R
Сила тока равна нулю в двух случаях:
1. либо электрический потенциал равен нулю,
2. либо существует очень большое сопротивление.
14. Механизм возникновения МПП
Движение специфических ионовчерез мембрану под действием
электрической силы может быть
только при одновременном
соблюдении двух условий:
1. мембрана содержит каналы, которые
проницаемы для данного вида ионов;
2. существует разность потенциалов по
обе стороны мембраны.
15. Ионный механизм мембранного потенциала покоя
Мембранный потенциал(МП) – это разность
потенциалов на мембране
нейрона, которую нейрон
имеет в данный момент
времени (Vm).
Мембранный потенциал
нейрона может быть
измерен с помощью
микроэлектрода,
помещенного в цитоплазму
нейрона и подсоединенного
к вольтметру.
16. Ионный механизм мембранного потенциала покоя
В состоянии покоя внутренняя сторона мембранызаряжена отрицательно, а внешняя сторона –
положительно.
Мембранный потенциал покоя (МПП) типичного
нейрона примерно равен - 65 mV.
Vm = -65 mV
Чтобы понять, каким образом возникает и
поддерживается МПП, необходимо рассмотреть
распределение некоторых ионов внутри нейрона и
окружающей его внешней среде.
17. Ионный механизм мембранного потенциала покоя
Равновесный потенциалРассмотрим гипотетическую клетку при
следующих условиях:
1. внутри клетки концентрация катионов K+ и
анионов А¯ выше, чем во внешней среде,
2. мембрана клетки не содержит ионных
каналов.
В этих условиях, несмотря на наличие
разности концентраций ионов,
1. не будет наблюдаться ток ионов через
мембрану;
2. мембранный потенциал будет равен
нулю.
18. Ионный механизм мембранного потенциала покоя
Ситуация изменится, если в мембране появятсяионные каналы, проницаемые для ионов K+, но
не проницаемые для анионов А¯.
Ионы K+ по градиенту концентрации начнут
перемещаться из клетки во внеклеточную среду.
За счет отрицательных ионов А¯ на внутренней
стороне мембраны начинает скапливаться
отрицательный заряд, а на внешней стороне
мембраны начинает появляться положительный
заряд.
Таким образом, на мембране нейрона начинает
появляться разность потенциалов.
19. Ионный механизм мембранного потенциала покоя
По мере увеличения разности потенциаловначинает возрастать электрическая сила,
которая толкает ионы K+ обратно в клетку (так
как положительно заряженные ионы K+
притягиваются к отрицательно зараженному
слою на внутренней стороне мембраны).
Когда на мембране достигается определенное
значение мембранного потенциала
электрическая сила, стремящаяся загнать
ионы K+ внутрь клетки, становится равной
химической силе градиента концентрации,
которая стремится вытолкнуть ионы K+ из
клетки.
Возникает состояние равновесия, при
котором сила электрической природы и сила
химической природы имеют одинаковое
значение, но направлены в разные стороны, а
движение ионов K+ приостанавливается.
20. Ионный механизм мембранного потенциала покоя
Ионный равновесный потенциал – это разностьпотенциалов на мембране, при которой сила химической и
электрической природы уравновешивают друг друга по
отношению к данному иону.
Например, калиевый равновесный потенциал равен
примерно – 80 mV.
Вывод: появление мембранного потенциала в нейроне
происходит автоматически при соблюдении двух условий:
1. существует разница концентраций ионов между внешней и
внутренней средой нейрона;
2. существует избирательная проницаемость мембраны
нейрона для данного иона.
21. Ионный механизм мембранного потенциала покоя
22. Ионный механизм мембранного потенциала покоя
Разница концентраций различных ионов вреальном нейроне
В реальном нейроне разные ионы по разному
распределены во внутриклеточной и внеклеточной
среде.
Ионы
Внеклеточная
концентрация
Внутриклеточная
концентрация
Отношение
Равновесный
потенциал
K+
5
100
1:20
-80 mV
Na+
150
15
10:1
62 mV
Ca2+
2
0,0002
10000:1
123 mV
Cl¯
150
13
11,5:1
-65 mV
23. Ионный механизм мембранного потенциала покоя
Каждый ион имеет свой собственныйравновесный потенциал.
Правило – концентрация ионов K+ больше
внутри клетки, а ионов Na+ и Cl¯ во
внешней среде.
Разница концентраций различных ионов
возникает в результате работы нескольких
ионных насосов, которые встроены в
мембрану нейрона.
24. Ионный механизм мембранного потенциала покоя
Два ионных насоса особенно важныдля понимания работы нейрона:
1. натрий-калиевый
2. кальциевый насос
Натрий-калиевый насос,
используя энергию АТФ, выкачивает
из клетки ионы Na+ и закачивает в
клетку ионы K+ против градиента
концентрации этих ионов.
За один цикл насос выкачивает
3
иона Na+ и 2 иона K+.
На работу этого насоса тратится
больше 70% всей АТФ,
находящейся в мозге.
25. Ионный механизм мембранного потенциала покоя
Кальциевый насос выкачивает из нейрона ионы Ca2+против градиента его концентрации.
1.
2.
Кроме того существуют дополнительные механизмы,
которые обеспечивают уменьшение концентрации ионов
Ca2+ в цитоплазме нейрона (0,00002 mM):
внутриклеточные белки, которые связывают данные
ионы;
клеточные органеллы (в частности, митохондрии и
эндоплазматический ретикулум), которые депонируют
(изолируют) ионы Ca2+.
26. Ионный механизм мембранного потенциала покоя
Значение ионных насосовБез ионных насосов в нейроне не смогла бы
поддерживаться разность концентрации
различных ионов, а, следовательно, в
нейроне не мог бы существовать
мембранный потенциал покоя, без которого, в
свою очередь, нейрон бы не смог отвечать на
внешнее воздействие и передавать
возбуждение.
27. Ионный механизм мембранного потенциала покоя
Относительная проницаемость мембраны для разных ионовВ реальном нейроне мембрана нейрона проницаема не для одного, а
для разных ионов.
Однако проницаемость мембраны для разных ионов разная.
Рассмотрим несколько сценариев для ионов Na+ и K+:
1. Если мембрана проницаема только для иона K+, то мембранный
потенциал будет равен калиевому равновесному потенциалу
(примерно -80 mV).
2. Если мембрана проницаема только для иона Na+, то мембранный
потенциал будет равен натриевому равновесному потенциалу
(примерно 62 mV).
3. Если мембрана имеет одинаковую проницаемость для ионов Na+ и K+, то
мембранный потенциал будет равен среднему значению между
натриевым и калиевым равновесным потенциалом (примерно - 9 mV).
28. Ионный механизм мембранного потенциала покоя
4. Если проницаемость мембраны в 40 раз больше для ионов K+, чемдля ионов Na+, то значение итогового мембранного потенциала
опять будет между натриевым и калиевым равновесным
потенциалом, но при этом ближе к калиевому равновесному
потенциалу.
Последний сценарий наиболее близок к ситуации в реальном
нейроне, в котором мембранный потенциал покоя равен -65 mV.
В реальном нейроне в состоянии покоя мембрана имеет высокую
проницаемость для ионов K+ и относительно низкую для ионов Na+.
29. Ионный механизм мембранного потенциала покоя
Вывод: высокая проницаемость мембранынейрона для ионов K+ является основным
источником мембранного потенциала
покоя (МПП), при этом относительная низкая
проницаемость мембраны для других ионов
(особенно ионов Na+) также вносит
определенный вклад в итоговое значение
МПП нейрона.
30. Ионный механизм мембранного потенциала покоя
Регуляция концентрации ионов K+ во внеклеточнойсреде
Мембранный потенциал очень чувствителен к
изменению концентрации ионов K+ во внеклеточной
среде. Например, если концентрация ионов K+ во
внешней среде уменьшится в 10 раз, то мембранный
потенциал покоя изменится от -65 до -17 mV.
Чувствительность мембранного потенциала к
концентрации ионов K+ привела в эволюции к
появлению механизмов, которые тонко регулируют
содержание этих ионов во внеклеточной среде:
1. гематоэнцефалический барьер
2. глиальные клетки (астроциты)
31. Ионный механизм мембранного потенциала покоя
Гематоэнцефалический барьер (ГЭБ) – этомеханизм, обеспечивающий ограниченный доступ
веществ, которые поступают через стенки капилляров,
к нейронам и глиальным клеткам внутри мозга.
Одна из функций ГЭБа – ограничение поступления из
крови ионов K+ во внеклеточную среду, окружающую
нейроны.
32. Ионный механизм мембранного потенциала покоя
Астроциты обеспечиваютрегуляцию концентрации
ионов K+ с помощью
калиевых насосов и
калиевых ионных каналов,
встроенных в их мембрану.
Когда внеклеточная
концентрация ионов K+
возрастает, эти ионы начинают
заходить внутрь астроцитов
через калиевые ионные
каналы.
33. Ионный механизм мембранного потенциала покоя
Вход ионов K+ в цитоплазмуастроцита приводит к повышению
локальной внутриклеточной
концентрации этих ионов,
которые начинают
распространяться по системе
разветвленных отростков в
другие части глиальной клетки.
Таким образом, астроциты
обладают глиальным
буферным механизмом,
который поддерживает
концентрацию ионов K+ во
внеклеточной среде на
постоянном уровне.
34. Ионный механизм мембранного потенциала покоя
ЗаключениеМеханизм возникновения МПП
1. Активность натрий-калиевого насоса обеспечивает и
поддерживает высокую концентрацию ионов K+ во
внутриклеточной среде нейрона.
2. Мембрана нейрона в состоянии покоя обладает высокой
проницаемостью для ионов K+, так как имеет многочисленные
калиевые каналы.
3. Движение ионов K+ через мембрану нейрона по градиенту их
концентрации приводит к появлению отрицательного заряда на
внутренней стороне мембраны и положительного заряда на
внешней стороне мембраны.
4. Разница потенциалов на мембране нейрона может
рассматриваться как заряд электрической батарейки, который
постоянно поддерживается за счет ионных насосов,
работающих на основе энергии АТФ.
Нейрон в отличие от других клеток способен возбуждаться. Под возбуждением нейрона понимают генерацию нейроном потенциала действия. Основная роль в возбуждении принадлежит другому типу ионных каналов, при открытии которых ионы натрия устремляются в клетку. Напомним, что благодаря постоянной работе насосных каналов концентрация натриевых ионов вне клетки примерно в 50 раз больше, чем в клетке, поэтому при открытии натриевых каналов ионы натрия устремляются в клетку, а ионы калия через открытые калиевые каналы начинают выходить из клетки. Для каждого типа ионов - натрия и калия - имеется свой собственный тип ионного канала. Движение ионов по этим каналам происходит по концентрационным градиентам, т.е. из места высокой концентрации в место с более низкой концентрацией.
Ответим на вопрос: как ионные каналы открываются и закрываются? В покоящемся нейроне натриевые каналы мембраны закрыты и на мембране, как это уже описывалось выше, регистрируется потенциал покоя порядка -70 мВ (отрицательность в цитоплазме). Если потенциал мембраны деполяризовать (уменьшить
Рис. 2.6. Работа натриевого потенциалзависимого канала.
А - канал открыт; Б - канал инактивирован; В - канал инактивирован и закрыт; 1 - мембрана; 2 - потенциалзависимый канал; 3 - инактивирующая частица.
поляризацию мембраны) примерно на 10 мВ, натриевый ионный канал открывается (рис. 2.6). Действительно, в канале имеется своеобразная заслонка, которая реагирует на потенциал мембраны, открывая этот канал при достижении потенциала определенной величины. Такой канал называется потенциалзависимым. Как только канал открывается, в цитоплазму нейрона устремляются из межклеточной среды ионы натрия, которых там примерно в 50 раз больше, чем в цитоплазме. Такое движение ионов является следствием простого физического закона: ионы движутся по концентрационному градиенту. Таким образом, в нейрон поступают ионы натрия, они заряжены положительно. Другими словами, через мембрану будет протекать входящий ток ионов натрия, который будет смещать потенциал мембраны в сторону деполяризации, т. е. уменьшать поляризацию мембраны. Чем больше ионов натрия войдет в цитоплазму нейрона, тем больше его мембрана деполяризуется. Потенциал на мембране будет увеличиваться, открывая все большее количество натриевых каналов. Но этот потенциал будет расти не бесконечно, а только до тех пор, пока не станет равным примерно +55 мВ. Этот потенциал соответствует присутствующим в нейроне и вне его концентрациям ионов натрия, поэтому его называют натриевым равновесным потенциалом. Вспомним, что в покое мембрана имела потенциал -70 мВ, тогда абсолютная амплитуда потенциала составит величину около 125 мВ. Мы говорим «около», «примерно» потому, что у клеток разного размера и типов этот
Рис. 2.7. Схема событий при возбуждении нейрона.
потенциал может несколько отличаться, что связано с формой этих клеток (например, количеством отростков), а также с особенностями их мембран.
Все изложенное выше можно формально описать следующим образом. В покое клетка ведет себя как «калиевый электрод», а при возбуждении - как «натриевый электрод». Однако после того как потенциал на мембране достигнет своего максимального значения +55 мВ, натриевый ионный канал со стороны, обращенной в цитоплазму, закупоривается специальной белковой молекулой. Это так называемая «натриевая инактивация» (см. рис. 2.6); она наступает примерно через 0,5-1 мс и не зависит от потенциала на мембране. Мембрана становится непроницаемой для натриевых ионов. Для того чтобы потенциал мембраны вернулся к исходному состоянию - состоянию покоя, необходимо, чтобы из клетки выходил ток положительных частиц. Такими частицами в нейронах являются ионы калия. Они начинают выходить через открытые калиевые каналы. Вспомните, что в клетке в состоянии покоя накапливаются ионы калия, поэтому при открывании калиевых каналов эти ионы покидают нейрон, возвращая мембранный потенциал к исходному уровню (уровню покоя). В результате этих процессов мембрана нейрона возвращается к состоянию покоя (-70 мВ) и нейрон готовится к следующему акту возбуждения.
Таким образом, выражением возбуждения нейрона является генерация на мембране нейрона потенциала действия. Его длительность
Рис. 2.8. Потенциал действия клетки желудочка сердца собаки.
в нервных клетках составляет величину около 1 / 1000 с (1 мс). Описанная последовательность событий приведена на рис. 2.7.
Подобные потенциалы действия могут возникать и в других клетках, назначение которых - возбуждаться и передавать это возбуждение другим клеткам. Например, сердечная мышца имеет в своем составе специальные мышечные волокна, обеспечивающие бесперебойную работу сердца в автоматическом режиме. В этих клетках также генерируются потенциалы действия (рис. 2.8). Однако они имеют затянутую, почти плоскую вершину, и длительность такого потенциала действия может затянуться до нескольких сот миллисекунд (сравните с 1 мс у нейрона). Такой характер потенциала действия мышечной клетки сердца физиологически оправдан, так как возбуждение сердечной мышцы должно быть длительным, чтобы кровь успела покинуть желудочек. С чем же связан такой затянутый потенциал действия у этого типа клетки? Оказалось, в мембране этих клеток натриевые ионные каналы не так быстро закрываются, как в нейронах, т. е. натриевая инактивация затянута.
Как ясно из этого описания, возбуждение (потенциал действия) нейрона сменяется так называемым «покоем». Однако никакого покоя в этот период нет. Как уже указывалось выше, в мембране есть еще и насосные каналы, количество которых примерно в 10 раз больше ионных, и они постоянно работают, откачивая из цитоплазмы излишек ионов натрия и закачивая туда недостающие ионы калия. Благодаря неустанной работе этих каналов нейрон всегда готов к возбуждению.
Описанный выше механизм возбуждения клетки (конечно, далеко не все клетки нашего организма способны возбуждаться) в основных чертах одинаков не только в нейронах и мышечных клетках человека, но и в аналогичных клетках других организмов. Например, в нейронах моллюсков, червей, крыс и обезьян при возбуждении происходят описанные выше последовательности событий. Более того, конструкция мембран, включая каналы, также примерно одинакова у всех организмов Земли.
Как уже указывалось, каналы представляют собой белковые молекулы, «прошивающие» мембрану (одна часть молекулы находится в цитоплазме, а другая - во внеклеточной среде). Интересно, что эти белковые молекулы, образующие ионный или насосный каналы, не вечны, а постоянно заменяются на новые (примерно каждые несколько часов). Все это свидетельствует об очень большой динамичности структуры нейрона.
Нейрон способен к возбуждению, которое состоит в том, что мембрана нейрона в состоянии покоя имеет потенциал порядка -70мВ (отрицательность β цитоплазме), а β состоянии возбуждения приобретает потенциал +55 мВ. Таким образом, абсолютная величина потенциала действия - около 125 мВ. Длительность потенциала действия нейрона составляет всего около 1 мс (1 / 1000 с).
Соединение в живых существах органической материи и металлов, использование живой клетки в качестве проводника электрического тока не могут не удивлять. Чем больше подробностей о своем теле я узнаю, тем чаще возникает удивление от простых его действий. Сгибание руки может восприниматься не как привычный, почти не замечаемый процесс, а как результат слаженной работы мышц и сухожилий, получивших команду от мозга, переданную по нервным волокнам с помощью электричества. Разглядывая полосу глубоко переливающейся синевы, возникающей на куске полированного лабрадорита, который я верчу в руках, я воспринимаю излучаемые им электрические волны рецепторами глаз. В рецепторах эта информация кодируется в виде череды электрических импульсов. Она передается в головной мозг, обрабатывается им, а результат всего этого — восприятие таинственно-красивого камня, возникающие при этом эмоции. Матрица, короче:)
Вместе с восхищением возникает и желание узнать — как это все происходит, желание вникать все глубже в подробности каждого процесса.
Потенциал действия — это тот электрический импульс, которой доставляет информацию от рецепторов к мозгу, и с помощью которого мозг управляет телом. Для его существования необходимы особо устроенные клетки, нейроны, мембраны которых облеплены с обеих сторон положительно и отрицательно заряженными ионами, которые только и ждут сигнала, чтобы этот электрический импульс пронести по всей длине клетки. Основным местом существования потенциала действия является аксон нейрона. Дендриты некоторых типов нейронов также способны проводить электрический импульс.

Достаточно взглянуть на картинку слева, чтоб увидеть основное отличие потенциала покоя от потенциала действия: когда мембрана находится в состоянии покоя, на всей ее протяженности снаружи существует положительный электрический заряд, а внутри — отрицательный.

А потенциал действия, проходящий по нейрону от тела клетки до конца аксона, представляет собой изменение внешнего и внутреннего зарядов клеточной мембраны на небольшом участке нейрона: на очень краткий промежуток времени отрицательный и положительный заряды по разным сторонам мембраны меняются местами.
Возникают вопросы: что запускает потенциал действия? Какова его физическая природа, что именно происходит в клетке при его существовании? Каким образом он распространяется? Почему он идет только в одном направлении? Как клетка восстанавливает исходный потенциал покоя после прохождения по ней волны возбуждения?
Итак, первый вопрос: как возникает потенциал действия? Так как передача сигналов от нейронов-источников к нейрону-получателю, или постсинаптическому нейрону (postsynaptic neuron ), происходит в синапсах, запускающий потенциал действия сигнал стоит искать именно там. Через синапс в нейрон приходят сигнальные вещества, но их поступление не обязательно означает запуск возбуждения нейрона. Если бы нейрон реагировал на очень малые количества сигнальных веществ, это было бы так же неудобно, как иметь супер-чувствительный выключатель, срабатывающий, если на него случайно чихнуть.

Необходимо достаточно сильное воздействие определенных сигнальных веществ, чтобы запустить потенциал действия. В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы — временные изменения зарядов мембраны, вызываемые пришедшими через синапс сигнальными веществами .
Постсинаптические потенциалы сильно отличаются от потенциала действия по своей природе. Если потенциал действия имеет одну строго определенную величину, то величина постсинаптических потенциалов зависит только от вида и количества вызвавших их химических веществ, переданных через синапсы. В то время как потенциалы действия, генерируемые разными нейронами, примерно одинаковы, постсинаптические потенциалы, возникающие в разных входных синапсах на одном и том же нейроне, сильно отличаются по величине и продолжительности. В одном синапсе нейрона приходящий нервный импульс может вызвать изменение мембранного потенциала на 0,1 мВ, а в другом — на 20 мВ. Кроме того, существуют как вызывающие потенциал действия, так и тормозящие его сигналы.
Тело клетки служит своеобразным калькулятором, простейшим аналоговым компьютером, суммирующим все поступающие в него сигналы в виде возникающих постсинаптических потенциалов. Аналоговые компьютеры устроены очень просто. Например можно построить водяной аналоговый компьютер. Если из одной трубы в банку вливается три килограмма воды, а из другой в ту же банку — пять, то взвесив банку и выяснив, что весы показывают вес, равный восьми килограммам, мы делаем вывод, что 3+5=8. На заре развития компьютерной техники использовались и водяные, и прочие аналоговые вычислительные машины, но наиболее продвинутой была, конечно, машина, которая в точности как и нейроны суммирует электрические токи. Операторы таких ЭВМ занимались вычислениями, перетыкая штекеры из одного гнезда в другое, сливая вместе разные токи.
Так вот в нейроне происходит такое же суммирование электрических токов. В процессе этого суммирования может быть перейден порог возбуждения (excitation threshold ): будет достигнуто определенное значение мембранного потенциала, которое запускает потенциал действия. Обрати внимание на то, что для достижения порога возбуждения мембранной потенциал должен увеличиваться. Если мембранный потенциал покоя нейрона равен -70 мВ, то его порог возбуждения может быть равен -55 мВ. Снижение мембранного потенциала например до -90 мВ приводит к тому, что для преодоления порога возбуждения требуется более сильное воздействие, ведь он остается прежним: -55мВ.

В постсинаптическом нейроне, получающем сигналы от двух возбуждающих и одного тормозящего нейронов, могут существовать различные комбинации сигналов, и далеко не каждая из них приводит к возникновению потенциала действия.
В первой схеме картинки вверху изображена ситуация, когда только от одного возбуждающего нейрона приходит слабый сигнал, которого недостаточно для преодоления порога возбуждения. Этот сигнал повторяется, но к моменту прихода второго, такого же слабого сигнала от того же аксона возбуждение мембраны уже достигает потенциала покоя, и потенциал действия не возникает.
На второй схеме изображен тот же самый слабый сигнал, но приходящий с большей частотой. Именно из-за увеличения частоты сигнала мембрана не успевает вернуться в состояние покоя в промежутке между двумя сигналами. В этом случае сумма остаточного заряда мембраны после первого сигнала и поступающего следом второго сигнала позволяет преодолеть порог возбуждения нейрона. В этом случае у основания аксона, которое называется аксонным холмиком (axon hillock [‘aksɒn ‘hɪlək] ), возникает потенциал действия.
На третьей схеме от обоих возбуждающих нейронов приходят слабые сигналы, и их суммы достаточно для запуска потенциала действия.
Четвертая схема показывает воздействие на постсинаптический нейрон возбуждающего и тормозящего сигнала. При поступлении только возбуждающего сигнала мембранный потенциал незначительно увеличивается, а при поступлении только тормозного сигнала — снижается. Если эти два сигнала приходят одновременно, то они всего лишь уничтожают друг друга, оставляя мембранный потенциал в состоянии, близком к потенциалу покоя. Это напоминает надавливание на одну и ту же дверь с разных сторон: по отдельности каждое воздействие могло бы приоткрыть дверь или наоборот, прикрыть ее, но существующие одновременно, они приводят лишь к небольшому сдвигу ее по направлению действия большей по модулю силы.
Природа системы такова, что даже малые по величине постсинаптические потенциалы, суммируясь, могут давать большой эффект. Нейрон «складывает» все поступившие в него возбуждающие постсинаптические потенциалы, «вычитает» поступившие тормозные постсинаптические потенциалы, и как только будет перейден порог возбуждения, возникает потенциал действия.
Чтобы разобраться, что происходит в клетке во время запуска и прохождения потенциала действия , необходимо перечислить каждого участвующего игрока и его действие, как было сделано при описании возникновения мембранного потенциала покоя в . Итак, действующие лица при создании потенциала действия :

1 . , которые были закрыты при существовании мембранного потенциала покоя, открываются, как только потенциал достигает порога возбуждения. Так как внутри клетки на мембране существует отрицательный заряд, то в этот момент через каналы туда врываются притягиваемые им
2 . положительно заряженные ионы натрия (Na +). Именно они создавали положительный заряд на внешней поверхности мембраны нейрона, и именно в этот момент они переносят его внутрь, оставляя на внешней мембране отрицательный заряд, который создают находящиеся вне клетки отрицательно заряженные ионы хлора.
3 . таким образом ионы хлора (Cl —) создают отрицательный заряд на внешней поверхности мембраны.

Перемещение ионов натрия внутрь клетки приводит к резкому изменению зарядов внутри и снаружи нейрона. Место существующего во время покоя нейрона отрицательного внутреннего заряда мембраны занимает положительный заряд ионов натрия. А снаружи клеточной мембраны после ухода оттуда Na + наконец-то может проявить себя Cl — , чей отрицательный заряд больше не компенсируется положительно заряженными ионами. Таким образом потенциал клеточной мембраны внезапно возрастает с -70 до +40 мВ. Именно это повышение, называемое деполяризацией (depolarization ), изображено резко идущей вверх частью пика на этой картинке. Такое повышение мембранного потенциала является сигналом закрыть потенциал-зависимые натриевые каналы, а также открывает потенциал-зависимые калиевые каналы, о которых раньше мы еще не говорили.
4 . потенциал-зависимые калиевые каналы. Они существуют в нейроне в дополнение к проточным калиевым каналам, которые, как ты наверное помнишь из прошлой главы про потенциал, не имеют ворот и обеспечивают постоянный отток калия из клетки, играя существенную роль в формировании потенциала покоя. Так вот оказывается, что есть две разновидности калиевых каналов, и сейчас мы рассмотрим работу потенциал-зависимого калиевого канала. Открываясь после того, как потенциал действия достиг своего пика, потенциал-зависимые калиевые каналы резко увеличивают отток ионов калия из клетки. Существующий в этот момент внутри нейрона положительный заряд, образовавшийся за счет притока Na + , активно выпихивает положительно заряженные ионы калия (К +) из клетки через оба вида каналов.

Итак, еще раз проследим цепочку событий до этого момента: потенциал достигает порога возбуждения, и этот электрический сигнал приводит к тому, что открылись потенциал-зависимые натриевые каналы, ионы Na + хлынули внутрь, притягиваемые отрицательно заряженными протеинами, произошла деполяризация, то есть разница зарядов снаружи и внутри резко снизилась, и даже более того — отсутствие положительно заряженных ионов натрия привело к возникновению отрицательного заряда там, где только что был положительный — произошла смена полюсов. Действие электрического импульса закончилось, натриевые каналы вновь закрылись, и столпившиеся внутри ионы Na + вытолкнули одноименно заряженные ионы К + через открывшиеся потенциал-зависимые калиевые каналы и через проточные калиевые каналы.
Заряд внутренней поверхности клеточной мембраны начинает стремительно снижаться, на схеме этому соответствует идущая вниз часть пика. Такое снижение заряда называется реполяризацией (repolarization ), то есть разница зарядов вне и внутри нейрона вновь восстановилась, как и было изначально в состоянии покоя. Но восстановилась именно разница зарядов, а не ионный состав, ее создающий. Несмотря на то, что в конце реполяризации снаружи мембраны образуется положительный заряд, равный положительному заряду мембраны в состоянии покоя, до достижения мембраной состояния покоя еще далеко. Ведь этот положительный заряд принесли туда вышедшие из клетки ионы К + , а при потенциале покоя его поддерживают ионы Na + , которые в данный момент заперты внутри клетки, в том самом месте, где ранее в состоянии покоя находились ионы калия.
В конце реполяризации может возникнуть снижение мембранного потенциала даже ниже его значения для покоящегося нейрона, или гиперполяризация (hyperpolarization [ˌhaɪpəpələraɪ’zeɪʃn] ). Через короткий промежуток времени она компенсируется дополнительным выходом ионов калия из клетки, и наступает прежнее, стабильное состояние покоя.

Когда мембранный потенциал становится равен потенциалу покоя (-70 мВ), потенциал-зависимые калиевые каналы закрываются. Через проточные калиевые каналы К + уже не вытекает так стремительно, ведь его концентрация внутри клетки упала, а снаружи находится много ионов калия. В то же время внутри находится много ионов натрия, которые должны быть выведены из клетки для того, чтоб через нее мог снова пройти потенциал действия.
Восстановлением исходного положения «натрий снаружи, калий внутри» занимается натрий-калиевый насос, выводящий из клетки три Na + и одновременно проносящий внутрь два К + . О его работе было рассказано в предыдущей главе про потенциал, и он схематично показан в четвертой части этой картинки. Совместные действия натрий-калиевого насоса и проточных калиевых каналов приводят к тому, что с внешней стороны мембраны опять возникает большая концентрация Na + , а концентрация К + внутри нейрона выше наружной.
Если бы работали только проточные калиевые каналы, то ионы калия только выходили бы из клетки, и было бы невозможным поддерживать внутри большую концентрацию К + , чем снаружи. Если бы действовали только потенциал-зависимые натриевые каналы, то ионы натрия вообще не могли бы покинуть клетку после того, как вошли туда, создавая потенциал действия. Но натрий-калиевый насос (ну то есть натрий-калиевый канал) решает обе эти проблемы: он выводит наружу Na + , восстанавливая его высокую концентрацию снаружи. И загоняет внутрь прошедшие через проточные каналы К + , не давая им возможности распределиться с обеих сторон мембраны равномерно, лишившись таким образом парциального давления, необходимого для формирования мембранного потенциала покоя.
Через тысячную долю секунды после того, как реполяризация нейронной мембраны достигла минимума, активная работа мембранных каналов восстанавливает потенциал покоя. Нейрон готов к проведению следующего потенциала действия.
5 . положительно заряженные ионы кальция (Ca 2+) играют важную роль в формировании потенциала действия лишь в некоторых нервных клетках, поэтому в данной главе, содержащей общую для различных нейронов информацию, она не будет описана.

На этом общее описание перемещения ионов через клеточную мембрану, превращающее потенциал покоя нейрона в потенциал действия, закончено. Можно сделать перерыв и переварить его вместе с чем-нибудь вкусным, заодно подпитав мозг, компенсировав те калории, которые он затратил на работу по обработке всей этой информации.
Распространение потенциала действия по всей длине аксона обеспечивается тем фактом, что возникший при его существовании в одном участке мембраны нейрона электрический ток возбуждает соседние участки, открывая натриевые потенциал-зависимые каналы и запуская уже там потенциал действия. Точнее, электрический ток запускает потенциал действия только с одной стороны от места существования предыдущего потенциала. Потому что с другой стороны натриевые каналы инактивированы после недавнего открытия, а без их участия возникновение потенциала действия невозможно. Если бы не было этого временного промежутка нефункциональности между двумя открытиями натриевых каналов, потенциал действия не мог бы распространяться только в одном направлении. Происходящая в результате цепная реакция открытия натриевых каналов обеспечивает передачу нервного импульса от места его возникновения до окончаний аксона, подходящих к другим нейронам (или к мышечной клетке).

Схематично эту цепную реакцию можно изобразить так:
Момент времени 1 : Электрический ток, возникший на в результате открытия потенциал-зависимых натриевых каналов на участке 1 , возбуждает соседние участки мембраны.
Момент времени 2 : В результате этого возбуждения на участке 2 открываются потенциал-зависимые натриевые каналы и возникает потенциал действия, который, в свою очередь, возбуждает соседние участки мембраны. На участке 1 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал после прохождения потенциала действия.
Момент времени 3 : Электрический ток, возникший в результате открытия потенциал-зависимых натриевых каналов на участке 2 , вызвал открытие таких же каналов на участке 3 , и потенциал действия перешел туда. На участке 1 потенциал-зависимые натриевые каналы не смогли открыться, несмотря на электрическое возбуждение от участка 2 , так как они были временно инактивированы после создания потенциала действия. На участке 2 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал покоя после прохождения потенциала действия.
Мы уже знаем, что отростки нейронов могут быть покрыты обернутыми вокруг них клетками нейроглии, или миелиновой оболочкой. Именно она делает аксон похожим на вытянутую цепочку сосисок, про ее образование и функции говорилось в . Передача потенциала возбуждения по «голым», не покрытым миелиновыми клетками, нервным волокнам отличается от его прохождения по миелинизированному аксону. При сравнении заметны существенные преимущества, предоставляемые обернутыми вокруг аксона клетками нейроглии:

1 . Экономичность . Мембрана аксона под миелином не имеет каналов для пропускания ионов, создающих потенциал действия. Каналы существуют только в узких промежутках между глиальными клетками, которые называют перехваты Ранвье (Ranvier’s constriction ), и только на этих узких участках аксона возникает потенциал действия. Концентрация ионных каналов в этих перехватах в 100 раз выше, чем в мембранах безмиелиновых волокон, тем не менее для того, чтобы обеспечить их работу, требуется значительно меньше энергии по сравнению с тем, сколько энергии потребовалось бы для обеспечения ионных каналов, расположенных по всей поверхности «голого» аксона. Кроме того, такая экономичность позволяет быстрее «перезаряжать» аксон для проведения следующего потенциала действия, поэтому миелинизированные волокна способны передавать сигналы более высокой частоты.
2
. Скорость
. По «голому» аксону сигнал передается в каждой его точке. Ионные каналы открываются в каждой точке его поверхности, один за другим, что напоминает тщательное, скрупулезное вчитывание в текст.
В миелинезированном аксоне потенциал действия, возникший в одном перехвате Ранвье, своим электрическим полем дотягивается до соседних перехватов, что приводит к возникновению в них новых потенциалов действия. То есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает следующий после поврежденного перехват, а в случае необходимости даже третий, четвертый и пятый по счету. Распространение электрического поля на такую длину возможно из-за электроизоляции, создаваемой миелиновыми муфтами: она уменьшает рассеивание электрического поля, которое неизбежно для безмиелиновых волокон.
Таким образом, скорость распространения потенциала действия по миелинизированным волокнам намного выше по сравнению с немиелинизированными. Ведь за один и тот же отрезок времени потенциал действия на «голом» аксоне возбудит лишь находящийся непосредственно рядом участок, а потенциал действия на миелинизированном волокне перепрыгнет на расстояние, равное расстоянию между одним или несколькими перехватами Ранвье:


Диапазон скоростей передачи потенциала действия в различных миелинизированных волокнах очень широк: от нескольких метров в секунду до «мирового рекорда», установленного аксоном креветки: он проводит возбуждение со скоростью, превышающей 200 м/с.
Чтобы представить, насколько эффективно может быть увеличена скорость проведения потенциала действия за счёт миелиновой оболочки, достаточно сравнить скорость его распространения по немиелинизированным и миелинизированным волокнам нейронов человека. Если взять скорость проведения импульса в одинаково тонких волокнах, то в безмиелиновом волокне она составит примерно 1 м/с, а в миелинизированном 15-20 м/с. В толстых аксонах человека, покрытых миелиновой оболочкой, скорость проведения может достигать 120 м/с.
Итак, подведем итог : передача электрического импульса через тело и отростки нейрона осуществляется с помощью прохождения ионов через клеточную мембрану.

1 . сначала на клеточной мембране создается потенциал покоя . Его образуют положительно заряженные ионы натрия (Na + ) снаружи клетки и отрицательно заряженные протеины внутри клетки. Создающие заряды частицы расположены очень близко к клеточной мембране, ведь они имеют разные знаки и притягиваются друг к другу. Поэтому потенциал покоя существует только возле мембраны, в целом среда внутри нейрона и вне его электрически нейтральна. Кроме Na + и отрицательно заряженных протеинов, в создании и поддержании электрического заряда участвуют положительно заряженные ионы (К + ) и отрицательно заряженные ионы хлора (Cl — ) , тоже находящиеся вблизи мембраны. Потенциал покоя равен примерно -70 мВ (в дальнейшем при указывании численного значения мембранных потенциалов слово «примерно» будет опускаться).
2 . когда нейрон через синапсы получает сигналы от других нейронов, под их влиянием на мембране тела нейрона начинают возникать постсинаптические потенциалы , то есть изменения величины электрических зарядов на внутренней и внешней поверхностях мембраны. Под влиянием этих сигналов внутренний отрицательный заряд и внешний положительный могут увеличиваться или уменьшаться на разные величины. И нейрон выступает в роли калькулятора, суммирующего эти воздействия.

3 . итогом суммирования постсинаптических потенциалов может являться продолжение нахождения нейрона в состоянии покоя. Но если сумма дойдет до значения, называемого порогом возбуждения , то в аксонном холмике (утолщении тела нейрона у основания аксона) возникнет потенциал действия .
4 . в создании потенциала действия главную роль играет клеточная мембрана . Она регулирует прохождение через нее ионов (всех перечисленных, кроме отрицательно заряженных протеинов, которые не могут покидать пределы клетки), открывая и закрывая специализированные каналы, способные проводить только определенные ионы.
5 . когда мембрана нейрона переходит порог возбуждения, в ней открываются потенциал-зависимые натриевые каналы . Исчезает то препятствие, которое отделяло положительно заряженные Na + от притягивающих их отрицательно заряженных протеинов, и Na + устремляются внутрь клетки . Таким образом наружный положительный заряд переносится внутрь клетки, а снаружи остается отрицательный заряд, создаваемый ионами Cl — . Происходит изменение мембранного потенциала от потенциала покоя, равного -70 мВ, до +40 мВ, и это называют потенциалом действия . Важно понимать, что это изменение потенциала происходит только на том участке мембраны, где открылись потенциал-зависимые натриевые каналы.
6 . В момент достижения мембраной потенциала в 40 мВ потенциал-зависимые натриевые каналы закрываются, и на этом же участке открываются потенциал-зависимые калиевые каналы . Снаружи в этот момент существует отрицательный заряд, а внутри — положительный, создаваемый вошедшими туда ионами Na + и уже находящимися там в большом количестве ионами К + . Ионы калия выпихиваются в окружающую нейрон среду через открывшиеся каналы, что резко снижает внутренний заряд мембраны и увеличивает внешний. По обеим сторонам мембраны создаются заряды той же величины, которые обычно поддерживают потенциал покоя. Но это состояние клетки отличается от потенциала покоя тем, что ионы калия и натрия находятся не на своих местах: Na + находится внутри, а К + снаружи. Опять же все описанное касается только одного места на мембране, в котором только что перестал существовать потенциал действия.
7 . Чтобы переместить положительно заряженные ионы Na + и К + по нужные для поддержания потенциала покоя стороны мембраны, используется натрий-калиевый насос . И это действие тоже касается ограниченного участка мембраны: того, где только что закрылись потенциал-зависимые калиевые каналы.
Один цикл работы натрий-калиевого насоса заключается в том, что он захватывает изнутри клетки три иона Na + , перемещает их наружу, где захватывает два иона К + и перемещает их внутрь . То есть канал обменивает три иона Na + на два иона К + , причем Na + могут перемещаться только наружу, а К + только внутрь клетки. Таким образом восстанавливается исходная концентрация натрия на наружной стороне мембраны и калия на внутренней. Натрий-калиевый насос выводит из клетки три положительно заряженных иона, а вводит в нее только два. Следовательно, при каждом его цикле внутренний положительный заряд мембраны уменьшается, а внешний увеличивается. К тому же и вытекающие ионы калия уносят изнутри наружу свои положительные заряды. Возникает вопрос: если потенциал мембраны уже и так был равен потенциалу покоя после входа ионов калия внутрь клетки, зачем его снижать еще сильнее? Ответ заключается в том, что кроме упомянутых здесь ионов, существуют и другие, чье участие в формировании потенциала покоя менее существенно, и именно для компенсации их влияния необходимо такое выравнивание мембранного потенциала. Итогом этого процесса является достижение мембранного потенциала покоя: заряд внутренней стороны мембраны отрицателен, внешней положителен, разность потенциалов равна -70 мВ.
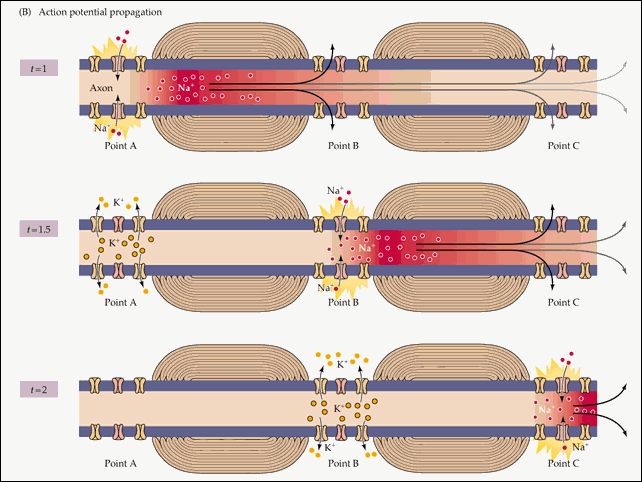
8 . Итак, в том месте клеточной мембраны, где всего три шага назад существовал потенциал действия, опять царит потенциал покоя. Но потенциал действия не прошел бесследно: его существование (описанное в пункте 5) создало электрическое поле, возбудившее соседние с ним части мембраны . С одной стороны (по направлению к концу аксона) это возбуждение вызвало открытие потенциал-зависимых натриевых каналов, запустив уже на новом участке потенциал действия. Который в свою очередь порождает электрическое поле, запускающее следующий потенциал действия, и этот цикл повторяется по всей длине аксона, от его основания до концов, просоединенных к следующим в нервной цепи клеткам.
С другой стороны от потенциала действия, ближе к телу нейрона, несмотря на воздействие электрического поля потенциал действия не возникает. Потому что потенциал-зависимые натриевые каналы на этом участке совсем недавно открывались, и на некоторое время они инактивированы. Именно эта пауза в их работе обеспечивает распространение потенциала действия только в одном направлении.
9 . Любой сигнал, будь то передача информации от сенсорного нейрона в мозг, или команда мозга, идущая к мотонейрону, передается одинаковыми по величине потенциалами действия . Результат потенциала действия (появление восприятия запаха, обусловленное работой сенсорных нейронов, или перемещение тела поближе к его источнику в результате слаженной работы мотонейронов) зависит исключительно от того, в какую нервную цепь включен нейрон, этот потенциал передающий. Для передачи интенсивности сигнала (силы запаха или величины сокращения мышцы) используется частота, с которой потенциалы действия передаются по нейрону. Слабый сигнал передается редкими, а сильный — частыми потенциалами действия .
10 . миелиновая оболочка вокруг отростков нейрона повышает скорость распространения потенциала действия за счет того, что сигнал передается на большое расстояние, равное ширине одной обернутой вокруг отростка клетки нейроглии, а не на непосредственно прилегающий к возбужденному участок, как это происходит у безмиелиновых волокон.




