Возникновение многоклеточных организмов. Как возникли многоклеточные организмы? В какую эру появились первые многоклеточные
Происхождение многоклеточных животных
Проблема происхождения многоклеточных животных представляет интерес не только для зоологии, но имеет большое общебиологическое значение. Многоклеточность представляет ту морфо-анатомическую основу, на которой формируется колоссальное разнообразие планов строения, жизненных форм и эволюционных потенций. Таким образом, знание путей и причин формирования многоклеточности у животных является ключом к пониманию многих важных зоологических и общебиологических вопросов.
В силу своего исключительного значения проблема происхождения многоклеточных животных издавна привлекала внимание исследователей. По этому поводу высказано немало гипотез, большинство их которых в настоящее время представляют исторический интерес, как любопытные примеры становления зоологической мысли. Все эти гипотезы группируются в четыре категории.
Первую группу составляют гипотезы, предполагающие независимое происхождение простейших и многоклеточных животных. К их числу относятся представления С. Авернцева (1910) и А.А. Заварзина (1945). По мнению этих авторов, еще на заре существования жизни на Земле первичное живое вещество (первородная слизь), еще не имевшее клеточной структуры, в одном случае приобрело организацию простейших, в другом – сразу многоклеточное строение. Подобные предположения противоречат как фундаментальному общебиологическому обобщению – клеточной теории, так и сравнительно-цитологическим данным, указывающим на исключительное сходство тонких клеточных структур Protozoa и Metazoa, которые едва ли могли возникнуть независимо.
Вторая группа представлена гипотезами, выводящими многоклеточных непосредственно от одиночных простейших. В частности, такой точки зрения придерживались Г. Иеринг (1877), А.А. Тихомиров (1887), И. Хаджи (1944), О. Штейнбок (1963) и другие авторы. Суть этих гипотез состоит в том, что многоклеточные животные произошли от крупных, высокоразвитых и сложноорганизованных простейших путем так называемой целлюляризации , т.е. единовременного разделения тела простейшего на множество специализированных клеток.
Подобное предположение, несмотря на всю его фантастичность с современных позиций, имеет определенные сравнительно-анатомические и эмбриологические основания. Так, некоторые инфузории по сложности организации, как минимум, не уступают низшим многоклеточным, таким, как бескишечным турбелляриям. Гипотезы целлюляризации исходят из того, что высокоспециализированные структуры инфузорий дали начало специализированным тканям и органам многоклеточных животных.
Эмбриологическим основанием гипотезы целлюляризации служит поверхностное дробление яиц членистоногих. При таком типе дробления деление ядер сперва не сопровождается делением цитоплазмы. Клеточные границы появляются одновременно и сравнительно поздно.
Гипотезы целлюляризации подверглись решительной критике со стороны В.А. Догеля, В.Н. Беклемишева, А.А. Захваткина, А.В. Иванова, О.М. Ивановой-Казас и других крупных зоологов. Суть этой критики, вкратце, состоит в следующем.
Во-первых, ее авторы указали на несостоятельность эмбриологического аргумента. Дело в том, что членистоногие – это животные, далеко отстоящие от истоков многоклеточности, и потому едва ли могли сохранить примитивные формы развития. Дробление яйца членистоногих, вне всякого сомнения – это результат далеко зашедшей специализации. Низшие же многоклеточные имеют совершенно иной ход онтогенеза.
Кроме того, исходя из гипотезы целлюляризации, в онтогенезе многоклеточного животного все ткани должны были бы дифференцироваться сразу после синцитиального дробления, и прямо на месте. В действительности же в ходе индивидуального развития многоклеточных (при гаструляции и органогенезе) наблюдаются последовательная дифференциация и значительные перемещения клеточных масс.
Во-вторых, высокоорганизованные простейшие – это слишком специализированные существа, чтобы дать начало животным с организацией принципиально иного типа. Подобное предположение противоречит одному из фундаментальных законов эволюции, гласящем, что у истоков эволюционно молодой группы организмов всегда находится не самый совершенный представитель из числа эволюционных предшественников.
В-третьих, гипотезы целлюляризации не имеют экологического обоснования. В этой связи, разделение тела простейшего на клетки выглядит беспричинным.
Третья группа гипотез выводит многоклеточных от колониальных простейших.
Среди них исторически первой явилась гипотеза гастреи знаменитого немецкого зоолога Эрнста Геккеля (1874), на длительный период завоевавшая большую популярность среди специалистов. В основу этой гипотезы Э. Геккель положил тот факт, что все многоклеточные животные в своем развитии обязательно проходят двуслойную стадию – гаструлу. Опираясь на биогенетический закон Геккеля-Мюллера (онтогенез есть краткое повторение филогенеза), Геккель предположил, что каждая стадия индивидуального развития многоклеточного животного повторяет (рекапитулирует) соответствующую стадию предковой формы. Так, стадии зиготы в филогенезе соответствует стадия одноклеточного организма, стадии морулы 1 (поздняя стадия дробления в виде плотного зачатка) отвечает морея – колония простейших в виде шаровидного скопления, стадии бластулы – колония простейших в виде полого шара, подобного современному Volvox – бластея . Впячивание (инвагинация) части стенки шаровидной колонии, по Геккелю, привело к образованию двуслойного животного – гастреи , отвечающего стадии инвагинационной гаструлы. Наружный слой клеток гастреи (эктодерма) выполнял функции кожного покрова, внутренний слой (энтодерма) играл роль кишечника. Бластопор выступал в качестве ротового отверстия (рис. 1). Среди современных Мetazoa ближе всего к гастрее Геккеля стоят наиболее примитивные представители кишечнополостных, которых Геккель положил в основу всего филогенетического древа многоклеточных животных.
Рис. 1. Происхождение многоклеточных животных по Геккелю.
Гипотеза гастреи Геккеля имела важное историческое значение, способствуя утверждению эволюционной идеи и крушению «теории типов» Ж. Кювье. Однако она была не свободна и от ряда принципиальных недостатков. К их числу относится, прежде всего, отсутствие сколь бы то ни было внятного экологического и физиологического обоснования процесса инвагинации.
Гипотеза гастреи Геккеля не осталась в одиночестве. Повинуясь зову моды, оригинальные гипотезы колониального происхождения многоклеточных животных предлагали многие ученые. Среди них я упомяну Ланкастера с его «теорией планулы» (1877) и Бючли с «теорией плакулы» (1884). Эти представления имеют в настоящее время лишь узкоспециальный исторический интерес, поэтому на них специально останавливаться не будем.
Весьма обстоятельную критику теории гастреи Геккеля дал И.И. Мечников (1886). Так, он убедительно продемонстрировал, что инвагинация не могла быть исторически первым способом образования двуслойной организации многоклеточных. Дело в том, что примитивным многоклеточным присущ совершенно другой механизм гаструляции, а именно мультиполярная иммиграция . Инвагинация же в филогенетическом аспекте появилась гораздо позднее, как результат прогрессивной эволюции онтогенеза многоклеточных. Кроме того, данные сравнительной физиологии однозначно указывают на вторичный характер полостного пищеварения, которому предшествовало внутриклеточное. Следовательно, по мнению И.И. Мечникова, едва ли первичные многоклеточные, подобно геккелевской гастрее, могли иметь кишечник и ротовое отверстие.
В качестве альтернативы гипотезе Геккеля, И.И. Мечников предложил оригинальную теорию, получившую название теории фагоцителлы (1886). Она достаточно хорошо обоснована и, в несколько модернизированном виде, сохраняет свое значение и в настоящее время.
При разработке теории фагоцителлы И.И. Мечников исходил из следующих соображений.
Предками многоклеточных животных могли быть одноклеточные с анимальным типом питания, то есть представители подцарства Protozoa.
Многие жгутиконосцы при определенных условиях (в частности, во время захвата пищи) могут принимать амебоидную форму.
Внутриклеточное пищеварение в ходе эволюции возникло раньше полостного, следовательно, первичные многоклеточные едва ли имели кишечник, равно как и ротовое отверстие.
Наиболее примитивными способами гаструляции выступают мультиполярная иммиграция и смешанная деламинация; филогенетические пути становления двуслойной организации должны были быть аналогичными.
Исходной стадией развития многоклеточных животных И.И. Мечников полагал шаровидную колонию жгутиконосцев, все особи которой располагались у ее поверхности одним слоем. Жгутики служили для плавания колонии и содействовали захвату пищевых частиц, организуя водовороты (так называемый седиментационный способ питания). Клетки, захватившие пищевую частицу, отбрасывали жгутик, принимали амебоидную форму и устремлялись внутрь колонии, где предавались перевариванию пищи. Завершив пищеварение и проголодавшись, они восстанавливали жгутик и возвращались на поверхность.
Впоследствии, по мнению И.И. Мечникова, исходно однородные особи колонии разделились на два пласта – кинобласт с локомоторной функцией, и фагоцитобласт с функцией трофической. Этот гипотетический организм напоминал личинок низших многоклеточных, известных под названием паренхимулы. Поэтому И.И. Мечников назвал это существо, по сложившейся зоологической традиции, паренхимеллой . Однако, имея в виду тот факт, что паренхимула является чисто расселительной стадией и самостоятельно не питается, передумал и предложил другое название – фагоцителла (делая тем самым акцент на внутриклеточном пищеварении этого животного).
Теория И.И. Мечникова, оказавшись в тени гипотезы гастреи Геккеля, должного признания не получила, а затем и вовсе была незаслуженно забыта. Лишь спустя полвека она была восстановлена в правах благодаря трудам В.Н. Беклемишева, А.В. Иванова, А.А. Захваткина и А.А. Заварзина. В настоящее время идеи И.И. Мечникова лежат в основе общепризнанных представлений о происхождении многоклеточных животных, о чем будет сказано ниже.
Четвертая группа гипотез предполагает происхождение многоклеточных животных от многоклеточных растений. Сформулировать подобные взгляды отважились Франц (1919, 1924) и Харди (1953).
Так, Франц предположил, что многоклеточные животные происходят от бурых водорослей, а именно от фукусов. Главные сходства, которым Франц придал абсолютное филогенетическое значение, заключаются в похожести жизненных циклов и более или менее одинаковом характере полового размножения.
Концепция Харди (1953) состоит в следующем. По мнению ее автора, переход к многоклеточному состоянию у растений протекает легче, чем у животных, поскольку общий характер питания многоклеточного растения – всасывание пищи всей поверхностью тела – сохраняется прежним. У животного же должен появиться новый способ питания, что чрезвычайно затрудняет переход к многоклеточному состоянию. В противном случае многоклеточность не дает животному никаких преимуществ.
Исходя из этих соображений, Харди предположил, что многоклеточные животные произошли от уже сформировавшихся метафитов, чем и преодолели вышеозначенные затруднения. Испытывая недостаток минерального питания, они начали питаться мелкими организмами, подобно тому, как это делают современные насекомоядные растения. В результате этих рассуждений у Харди появился простой полипообразный метазоон с пузыревидной полостью и щупальцами.
Гипотезы происхождения метазоев от метафитов настолько экзотичны, что останавливаться на их критическом анализе нет смысла.
Современные представления о происхождении многоклеточных животных
Современные представления о происхождении многоклеточных животных основаны на гипотезе фагоцителлы И.И. Мечникова, несколько модернизированной и дополненной с учетом более поздних открытий и идей.
Прежде чем попытаться реконструировать ход этого процесса, следовало бы задуматься: а зачем, собственно говоря, эта самая многоклеточность животным вдруг понадобилась? Существовали они на Земле миллиард лет, совершенствуясь в рамках одноклеточной организации, и вдруг занялись созданием «государства клеток»?
Специалистам хорошо известно, что всякое развитие выходит на качественно новый уровень тогда и только тогда, когда исчерпываются возможности развития в рамках старого качества. Другими словами, когда развитие упирается в некий «потолок», который не может быть преодолен на основе прежней организации. Это означает, что одноклеточное существо обладает некими принципиальными ограничениями, которые мешают ему совершенствоваться.
Анализ зоологического материала позволил установить, что к числу таких ограничений относятся, прежде всего, некоторые аллометрические зависимости. Известно, что эволюция жизни на Земле идет по пути усложнения, одним из проявлений которого является так называемый филогенетический рост – последовательное увеличение размеров организмов по мере их филогенетического развития.
У одноклеточных этот рост связан со многими факторами. Прежде всего, с необходимостью движения простейшего относительно среды. Дело в том, что снабжение простейших кислородом и удаление продуктов их жизнедеятельности происходит путем диффузии. В результате, одноклеточное существо очень быстро создает вокруг себя «пустыню», к тому же загаженную собственными выделениями. Поэтому для него жизненно важно сменить обстановку, т.е. переместиться из испорченного пункта «А» в свежий пункт «Б». Однако мелкий организм имеет колоссальное отношение поверхности к объему и, в силу этого, очень страдает от трения о воду. Другими словами, при активном движении он испытывает исключительно большое сопротивление среды. Причем это сопротивление пропорционально площади простейшего, а его локомоторная мощность – объему. Таким образом, увеличение линейных размеров простейшего, скажем, вдвое приведет к тому, что сопротивление плаванию возрастет в четыре раза, а мощность – в восемь раз. Или, что то же самое, удельная мощность (отношение мощности к силам трения) увеличится в два раза! В результате, возникает тенденция на увеличение размеров тела простейшего как эволюционный ответ на потребность в энергичном плавании.
Другая причина увеличения размеров – создавать в своем теле запас питательных веществ и резервной биомассы, что делает их относительно независимыми от колебаний жизненных ресурсов.
И, наконец, третья (но не последняя!) причина – филогенетический рост есть простое следствие усложнения организации. Когда различных морфологических структур много, им необходим более вместительный «контейнер».
Таким образом, в ходе прогрессивной эволюции простейшие обязаны увеличиваться в размерах. Может ли этот процесс идти бесконечно? И почему мы не вправе ожидать появления, скажем, сложно устроенного одноклеточного величиной со слона?
Дело в том, что функционирование одноклеточного существа, как и любого другого живого организма, основано на целесообразных реакциях на вызовы среды. У одноклеточных такие реакции управляются со стороны ядра. Например, в среде появилось некое вещество. Это вещество вступает в связь с рецепторами, находящимися на внешней поверхности клеточной мембраны, и в результате этого взаимодействия рецептор посылает ядру химический сигнал в виде какой-либо молекулы. Эта молекула достигает ядра и вызывает экспрессию нужного гена. В результате, клетка начинает синтезировать нужное вещество: ответ состоялся.
При увеличении размеров простейшего расстояние между клеточной мембраной и ядром увеличивается. Увеличивается и время реакции организма на внешние сигналы, и он, в конце концов, начинает безнадежно запаздывать, уподобляясь очень флегматичному и неповоротливому великану, беззащитному в быстро меняющейся обстановке.
Надо сказать, что с подобной проблемой живая природа сталкивалась не один раз. Сравнительно недавний пример: появление на Земле крупных динозавров, имеющих длину от носа до кончика хвоста свыше 20 метров. Учитывая, что скорость проведения нервного импульса у рептилий составляет величину того же порядка (30-40 метров в секунду), можно себе представить, как маленький, но дерзкий хищник изволил отобедать хвостом динозавра прежде, чем тот начал осознавать, что у него в тылу что-то происходит. Существует мнение, что именно в силу этих причин головной мозг гигантов был не крупнее теннисного мячика, тогда как основной объем нервной массы помещался в крестцовом отделе. Это «изобретение» уменьшало «плечо» рефлекторной дуги примерно вдвое, во столько же раз сокращая время «осмысления» динозавром тех событий, которые происходят с его хвостом.
Какой же выход из создавшейся ситуации нашли простейшие? Этот выход состоял в полиэнергидности: простейшее развило множество ядер, каждое из которых управляло собственной «провинцией» – прилегающим участком цитоплазмы.
Однако и этот выход оказался лишь полумерой, поскольку целостность получившегося организма была невелика. Простейшее разделилось на множество «автономий», а его координированное управление как единого целого затруднялась все теми же расстояниями между клеточной мембраной и глубоко лежащими частями клетки. В этом отношении простейшее уподобилось огромной и неповоротливой Российской империи середины XIX века, когда приказы из столицы, каковой в те времена был Петербург, передавались в удаленные губернии по конной эстафете. Стоит ли удивляться, что при такой постановке дела губернатор Камчатки узнал Крымской войне лишь спустя три месяца после ее начала, да и то это известие он получил не из Петербурга, а от английской эскадры, приступившей к бомбардировке Петропавловска-Камчатского с морского рейда.
Таким образом, общий план строения простейшего оказался чреват принципиальными ограничениями, которые в рамках одноклеточной организации не могут быть преодолены. Тем не менее, магистральная линия эволюции простейших от примитивных одноядерных диплоидных форм к полиплоидным и, далее, полиэнергидным представителям подцарства совершенно справедливо интерпретируется зоологами как тенденция к многоклеточности .
Коль скоро существуют проблемы, которые не получается разрешить в рамках одноклеточной организации, остается один путь – кооперация одноклеточных индивидов. Именно эти соображения лежат в основе современных представлений о возникновении многоклетчности у животных.
Согласно этим представлениям, предками многоклеточных животных явились довольно примитивные жгутиконосцы, сходные с современными представителями Choanophlagellata – воротничковыми жгутиконосцами. На их филогенетическую близость указывают сходства в ультратонком строении жгутика и кинетосомы, митохондрий, составе запасных питательных веществ, а также наличие воротничковых клеток или клеток с рудиментами воротничка в составе некоторых многоклеточных животных. К тому же, современные Choanophlagellata демонстрируют четко выраженную склонность к образованию различных колоний.
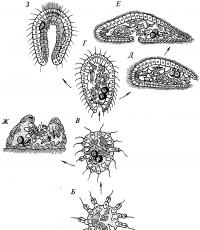
Первой стадией на пути к многоклеточности явилось объединение одиночных воротничковых жгутиконосцев в просто устроенную колонию типа Sphaeroeca – шаровидный агрегат клеток, ориентированных своими жгутиками наружу (рис. 2). Клетки колонии были совершенно одинаковыми в морфологическом и функциональном отношениях. Самое большее, на что могла быть способна такая колония в плане дифференциации клеток, это возникновения морфологического градиента, как это имеет место у современного Volvox – на функционально переднем его полюсе клетки мельче, и постепенно увеличиваются по направлению к функционально заднему полюсу.

Рис. 2. Происхождение многоклеточных животных по Иванову.
Что побудило одноклеточных индивидов к объединению в колонию? По-видимому, необходимость преодоления тех самых аллометрических ограничений, которые мешают простейшим плавать. Поверхность шаровидной колонии и, следовательно, ее трение о воду значительно меньше, чем совокупная поверхность составляющих ее индивидов, а локомоторная мощность колонии равна сумме локомоторных мощностей индивидов. Таким образом, кооперация простейших повышала эффективность плавания.
Сначала такая колония размножалась, по-видимому, только бесполым путем, распадаясь на отдельные клетки, каждая из которых давала начало новой колонии (так, как это происходит у современных Sphaeroeca ). Для того чтобы колония могла развиваться как единое целое, должна была произойти первая дифференцировка клеток на половые и соматические. Точнее, в цикле развития предковой формы должно было появиться новое поколение, представленное половыми особями, подобно тому, как это наблюдается в колониях современных Volvox или Proterospongia. Специализированные половые клетки спасали колонию от постоянного разрушения, так как брали функцию размножения на себя. Колония получила возможность прогрессировать как единое образование, ее интеграция могла усиливаться от поколения к поколению и подчинять себе индивидуальность отдельных особей.
Вторая стадия – факультативная дифференциация колонии на функциональные группы клеток. Причина возникновения этой стадии – продолжающееся увеличение размеров колонии, в силу чего составляющие ее клетки разошлись по периферии, а внутри образовалось свободное пространство, заполненное студенистой массой. Поскольку жгутиковые клетки располагались на поверхности колонии, локомоторная мощность стала пропорциональной площади, и дальнейшее совершенствование локомоции за счет простого увеличения размеров оказалось невозможным – в данном отношении эволюция зашла в тупик. Зато у членов колонии появилась возможность чередовать разные фазы активности, оптимизируя выполнение той или иной функции поочередно. Так, клетки, находящиеся снаружи, выполняли локомоторную функцию в интересах всей колонии, и питались поодиночке, отфильтровывая из воды пищевые частицы, каждый для себя. «Нагрузившись» пищей, клетки утрачивали жгутик, приобретали амебоидную форму и уходили внутрь колонии, где сосредоточивались на переваривании пищи. Опять проголодавшись, клетки возвращались на поверхность, восстанавливали жгутик, и все начиналось сначала.
Третья стадия . Такое физиологическое обособление клеточных слоев явилось важной предпосылкой для постоянной морфологической дифференциации колонии. В конце концов, клеточная масса колонии подразделилась на два пласта, каждый из которых специализировался на выполнении тех или иных функций. Клетки наружного слоя – кинобласт – приняли на себя функцию локомоции и, частично, захвата пищи (с последующей ее передачей клеткам внутреннего слоя). Клетки внутреннего слоя – фагоцитобласт – приняли на себя трофическую функцию – захвата пищи с поверхности колонии и ее переваривания (с последующей передачей легко усваиваемых продуктов переваривания клеткам кинобласта). Таким образом, клеткам не надо было больше сменять друг друга в своем движении с поверхности колонии внутрь и обратно, меняя при этом облик жгутиконосца на форму амебы и форму амебы на облик жгутиконосца.
Таким образом, постоянная дифференциация членов колонии на два клеточных пласта позволила экономить время и жизненные ресурсы членов колонии, оказалась выгодной в эволюционном отношении и закрепилась генетически. Так возникло первое первичное многоклеточное животное (Prometazoa), получившее название ранняя фагоцителла , или фагоцителла -1.
Четвертая стадия – появление первого истинного многоклеточного животного Eumetazoa. Суть ее состояла в эпителизации кинобласта, повлекшей за собой серию важных эволюционных последствий. Сама же эпителизация была вызвана, в первую очередь, необходимостью повышения прочности межклеточных соединений достаточно крупного и активно плавающего существа. Таким образом, клетки кинобласта оказались надежно «сшиты» друг с другом и образовали самую первую ткань – эктодерму .
Это повлекло за собой следующие преобразования.
1. При эпителизации кинобласта часть клеток специализировалась на чувствительной функции и функции проведения раздражения. Так возникли первые чувствительно-нервные элементы, образующие в эктодерме первичный нервный плексус , или нервную систему диффузного типа. Чувствительные элементы сконцентрировались на аборальном полюсе, где образовали теменную пластинку . В конце концов, у фагоцителлы очень рано возник координационный центр, на основе которого развился первичный мозг (возможно, ассоциированный со статоцистом). Благодаря этому интегрированность фагоцителлы как целостного организма резко возросла.
2. Клетки фагоцитобласта больше уже не могли просовывать свои псевдоподии между жгутиковыми клетками, поэтому в эктодерме появилось отверстие – бластопор, или первичный рот, через которое клетки фагоцитобласта могли захватывать пищевые частицы. Ротовое отверстие возникло на функционально заднем полюсе, поскольку при плавании фагоцителлы, в силу гидродинамических причин, именно там концентрировались пищевые частицы. Последнее доказывается лабораторными опытами с личинками некоторых низших многоклеточных животных, рекапитулирующих позднюю фагоцителлу: при добавлении в воду частиц туши все они оказываются в области заднего полюса личинки, где и фагоцитируются клетками пищеварительной паренхимы.
3. С появлением ротового отверстия функционально задний полюс стал и морфологически задним, и получил название орального (или вегетативного). В соответствие с этим, противоположный полюс стал называться аборальным (или анимальным), а соединяющая их ось – первичной главной осью тела , очень важной координатой, с которой при морфоанатомическом анализе соотносится топографическое положение органов и частей всех многоклеточных животных. Таким образом, возникло существо, обладающее радиальной гетерополярной симметрией – первичной формой симметрии настоящих многоклеточных.
Это существо получило название поздней фагоцителлы , или фагоцителлы-2 . Именно оно лежит в основе филогенетического древа всех истинных многоклеточных животных Eumetazoa.
1 Морула в переводе с латинского означает тутовая ягода.
На Земле лишь самые простые существа состоят из одной клетки. Все сложно организованные растения, животные и грибы состоят из нескольких клеток, и у большинства многоклеточных организмов клеток действительно очень много.Переход к дыханию кислородом вызвал необходимость того, чтобы весь кислород доходил до всех клеток. Но вначале кислорода было довольно мало, так что его было недостаточно для проникновения в глубь клеточных слоев. И лишь когда деятельность фотосинтезирующих одноклеточных привела к тому, что атмосфера Земли насытилась кислородом, появились многоклеточные организмы.
Узнать, какова была концентрация кислорода в прошлом, позволяют геологические изыскания. Некоторые минералы не могут существовать в атмосфере, богатой кислородом, и, если бы их удалось обнаружить в каком-нибудь слое земной коры, то это означало бы, что кислорода в те времена было довольно мало. Хотя жизнь возникла довольно давно, первые многоклеточные существа появились чуть более одного миллиарда лет назад. Это были растения.
Многоклеточные животные появились еще позже - 600 миллионов лет назад. Как ни странно, это были крупные существа, напоми-
Пейзаж времен палеозоя
нающие медуз. В те времена на всей планете не было ни одного хищника.
По всей видимости, первые многоклеточные существа не стали предками современных многоклеточных организмов, видимо, многоклеточные существа возникали не один раз. Первые существа с твердым скелетом появились около 540 миллионов лет назад. Об этих организмах мы знаем гораздо больше, их облик известен нам гораздо лучше, чем внешний вид самых древних организмов, ведь по скелету -
неважно, раковине, панцирю или кости - можно представить то существо, чьей частью он был.
Поскольку до момента появления скелета ясных отпечатков не оставалось, все, что происходило до этого, назвали эрой скрытой жизни, или криптозоем, а все, что произошло потом, - палеозоем. Скелет стал настоящей революцией. Это опора, а значит движение, высокий рост, защита, возможность противостоять силе тяжести на суше и завоевать новые пространства.
Нужно помнить, что суша в то время была безжизненной, и все живое существовало лишь в океане. Что же вызвало появление скелета у древних организмов? Вероятно, увеличение количества кислорода позволило вести более активную жизнь. Активность привела к быстрому накоплению твердых отходов в организме и развитию скелета.
Некоторые организмы образовывали колонии таких размеров, что меняли облик древних водоемов. Это известковые водоросли и губки.
Вероятно, первыми существами, которые начали уничтожать других, стали головоногие моллюски, к которым относятся современные осьминоги, каракатицы и кальмары.
Несмотря на то что позвоночные животные появились тоже достаточно давно, они занимали не самые почетные места в древнем раскладе сил. Бесчелюстные панцирные - пред
ки рыб, уже были похожи на рыб, которых мы едим или разводим в аквариумах. Панцирь покрывал и бесчелюстных, и первых настоящих рыб. Но рыбы обрели привычный для нас облик лишь миллионы лет спустя.
Еще по теме КАК ВОЗНИКЛИ МНОГОКЛЕТОЧНЫЕ ОРГАНИЗМЫ?:
- КАК ВОЗНИКЛА ИДЕЯ БИЗНЕСА И СОУЧРЕДИТЕЛИ ПОЗНАКОМИЛИСЬ ДРУГ С ДРУГОМ
- 6. Поздний докембрий: возникновение многоклеточности. Гипотеза кислородного контроля. Эдиакарский эксперимент.
Все живые организмы разделяются на подцарства многоклеточных и одноклеточных существ. Последние представляют собой одну клетку и относятся к простейшими, в то время как растения и животные являются теми структурами, в которых веками развивалась более сложная организация. Количество клеток варьируется в зависимости от разновидности, к которой относится особь. Размер большинства настолько мал, что увидеть их можно только под микроскопом. Клетки появились на Земле примерно 3,5 миллиарда лет назад.
В наше время все процессы, происходящие с живыми организмами, изучает биология. Подцарством многоклеточных и одноклеточных занимается именно эта наука.
Одноклеточные организмы
Одноклеточность определяется наличием в организме единственной клетки, которая выполняет все жизненные функции. Всем известные амеба и инфузория-туфелька представляют собой примитивные и, вместе с тем, древнейшие формы жизни, которые являются представителями этого вида. Они были первыми живыми существами, что обитали на Земле. Сюда же входят такие группы, как споровики, саркодовые и бактерии. Все они малы и в основном невидимы для невооруженного глаза. Их принято разделять на две общие категории: прокариотические и эукариотические.
Прокариоты представлены простейшими или грибами некоторых видов. Кто-то из них живет колониями, где все особи одинаковы. Весь процесс жизни осуществляется в каждой отдельной клетке для того, чтобы она выжила.
Прокариотические организмы не имеют связанных мембранами ядер и клеточных органелл. Это обычно бактерии и цианобактерии, такие как кишечная палочка, сальмонеллы, ностоки и др.
Все представители этих групп различаются по размеру. Самая малая бактерия имеет длину всего 300 нанометров. Одноклеточные обычно обладают специальными жгутиками или ресничками, которые участвуют в их передвижении. Они имеют простое тело с выраженными основными чертами. Питание, как правило, происходит в процессе поглощения (фагоцитоза) пищи и хранится в специальных органоидах клетки.
Одноклеточные доминировали как форма жизни на Земле в течение миллиардов лет. Однако эволюция от простейших к более сложным особям изменила весь ландшафт, поскольку она привела к зарождению биологически развитых связей. Кроме того, появление новых видов привело к образованию новой среды с разнообразными экологическими взаимодействиями.
Многоклеточные организмы
Основной характеристикой подцарства многоклеточных является наличие в одном индивидууме большого количества клеток. Они скрепляются между собой, тем самым создавая совершенно новую организацию, которая состоит из множества производных частей. Основное количество из них можно увидеть без каких-то специальных приборов. Растения, рыбы, птицы и животные выходят из единственной клетки. Все существа, входящие в подцарство многоклеточных, регенерируют новые особи из зародышей, которые формируются из двух противоположных гамет.
Любая часть особи или цельный организм, который определяется большим количеством составляющих, является сложной, высоко развитой структурой. В подцарстве многоклеточных классификация четко разделяет функции, при которых каждая из отдельных частиц выполняет свою задачу. Они занимаются процессами жизнедеятельности, поддерживая этим существование всего организма.
Подцарство Многоклеточные на латыни звучит как Metazoa. Чтобы сформировать сложный организм, клетки нужно идентифицировать и присоединить к другим. Только с десяток простейших можно заметить индивидуально невооруженным глазом. Остальные почти два миллиона видимых особей являются многоклеточными.
Плюрицеллюлярные животные созданы результатом объединения особей путем образования колоний, нитей или агрегации. Плюрицеллюлярные развивались самостоятельно, вроде вольвокса и некоторых жгутиковых зеленых водорослей.
Признаком подцарства многоклеточных, то есть его ранних примитивных видов, было отсутствие костей, раковин и других твердых частей тела. Поэтому их следов не сохранилось до наших дней. Исключением являются губки, обитающие в морях и океанах до сих пор. Возможно, их останки находятся в каких-нибудь древних скалах, как, например, Grypania spiralis, окаменелости которых найдены в древнейших слоях черного сланца, относящегося к раннепротерозойской эре.
В находящейся ниже таблице подцарство многоклеточных представлено во всем его многообразии.

Сложные взаимосвязи возникли в результате эволюции простейших и появления способности клеток разделяться по группам и организовывать ткани и органы. Существует много теорий, объясняющих механизмы, с помощью которых одноклеточные могли эволюционировать.
Теории возникновения
На сегодняшний день существуют три основных теории возникновения подцарства многоклеточных. Краткое содержание синцитиальной теории, чтобы не углубляться в подробности, можно описать в нескольких словах. Суть ее состоит в том, что примитивный организм, который имел в своих клетках несколько ядер, мог со временем разделить внутренней мембраной каждое из них. Например, несколько ядер содержит грибок плесени, а также инфузория-туфелька, чем подтверждают эту теорию. Однако наличия нескольких ядер недостаточно для науки. Чтобы подтвердить теорию их множественности, необходимо наглядное превращение в хорошо развитое животное простейшего эукариота.
Теория колоний говорит, что симбиоз, состоящий из разных организмов одного вида, привел к их изменению и появлению более совершенных существ. Геккель — первый ученый, кто представил эту теорию в 1874 году. Сложность организации возникает потому, что клетки остаются вместе, а не разъединяются в процессе деления. Примеры этой теории можно увидеть у таких простейших многоклеточных, как зеленые водоросли, которые называются эвдорина или вольвакса. Они образуют колонии, которые насчитывает до 50000 клеток в зависимости от вида.
Теория колоний предлагает слияние различных организмов одного вида. Преимущество этой теории заключается в том, что было замечено, как во время нехватки продовольствия амебы группируются в колонию, которая передвигается словно единое целое, в новое место. Какие-то из этих амеб немного отличаются друг от друга.
Однако проблема этой теории заключается в том, что неизвестно, как ДНК разных особей могут быть включены в единый геном.
Например, митохондрии и хлоропласты могут быть эндосимбионтами (организмами в организме). Это случается крайне редко, и даже тогда геномы эндосимбионтов сохраняют между собой различия. Они отдельно синхронизируют свою ДНК во время митоза видов хозяев.
Два или три симбиотических индивидуума, образующих лишайник, хотя и зависят друг от друга ради выживания, но должны отдельно размножаться, а затем повторно соединяться, снова создавая единый организм.
Другие теории, которые также рассматривают возникновение подцарства многоклеточных:
- Теория GK-PID. Около 800 миллионов лет назад незначительное генетическое изменение в одной молекуле под названием GK-PID, возможно, позволило особям перейти от одной клетки к более сложной структуре строения.
- Роль вирусов. Недавно было признано, что гены, позаимствованные у вирусов, играют решающую роль в делении тканей, органов и даже при половом размножении, при слиянии яйцеклетки и сперматозоида. Был найден первый белок syncytin-1, который передался от вируса к человеку. Он находится в межклеточных мембранах, которые разделяют плаценту и мозг. Второй белок был выявлен в 2007 году и назван EFF1. Он помогает формировать кожу круглых червей нематод и является частью целого семейства белков FF. Доктор Феликс Рей в Институте Пастера в Париже построил 3D-макет структуры EFF1 и показал, что это он связывает частицы вместе. Этот опыт подтверждает тот факт, что все известные слияния мельчайших частиц в молекулы имеют вирусное происхождение. Это также говорит о том, что вирусы были жизненно важны для коммуникации внутренних структур, и без них было бы невозможным появления колонии подцарства многоклеточных типа губок.
Все эти теории, как и многие другие, которые продолжают предлагать известные ученые, очень интересны. Однако ни одна из них не может четко и однозначно ответить на вопрос: как из единственной клетки, которая зародилась на Земле, могло появиться такое огромное разнообразие видов? Или: почему одиночные особи решили объединиться и стали существовать вместе?
Может, пройдет несколько лет, и новые открытия смогут нам дать ответы на каждый из этих вопросов.

Органы и ткани
Сложные организмы имеют такие биологические функции, как защита, кровообращение, пищеварение, дыхание и половое размножение. Они выполняются определенными органами, такими как кожа, сердце, желудок, легкие и половая система. Они состоят из множества различных типов клеток, которые работают сообща для выполнения конкретных задач.
Например, сердечная мышца имеет большое количество митохондрий. Они производят аденозинтрифосфат, благодаря которому кровь беспрерывно движется по кровеносной системе. У клеток кожи, наоборот, меньше митохондрий. Вместо этого они имеют плотные белки и производят кератин, который защищает мягкие внутренние ткани от повреждений и внешних факторов.
Размножение
В то время как все без исключения простейшие организмы размножаются бесполым путем, многие из подцарства многоклеточных предпочитают половое размножение. Люди, например, являются сложнейшей структурой, созданной путем слияния двух одиночных клеток, называемых яйцеклеткой и сперматозоидом. Слияние одной яйцеклетки с гаметой (гаметы - это специальные половые клетки, содержащие один набор хромосом) сперматозоида приводит к образованию зиготы.
Зигота содержит генетический материал как спермы, так и яйцеклетки. Деление ее приводит к развитию абсолютно нового, отдельного организма. Во время развития и деления клетки, согласно заложенной в генах программе, начинают дифференцироваться по группам. Это в дальнейшем позволит им выполнять совершенно разные функции, несмотря на то что они генетически идентичны друг другу.
Таким образом, все органы и ткани организма, которые образуют нервы, кости, мышцы, сухожилия, кровь, — все они возникли из одной зиготы, появившейся благодаря слиянию двух одиночных гамет.
Преимущество многоклеточных
Есть несколько основных преимуществ подцарства многоклеточных организмов, благодаря которым они доминируют на нашей планете.
Поскольку сложное внутреннее строение позволяет увеличить размер, оно также помогает развивать структуры и ткани более высокого порядка с многочисленными функциями.
Крупные организмы имеют лучшую защиту от хищников. Они также обладают большей мобильностью, что позволяет им мигрировать в более благоприятные для проживания места.
Есть еще одно неоспоримое преимущество подцарства многоклеточных. Общая характеристика всех его видов — это достаточно долгая продолжительность жизни. Тело клетки подвергается воздействию окружающей среды со всех сторон, и любое ее повреждение может привести к гибели индивидуума. Многоклеточный организм будет продолжать существовать, даже если одна клетка погибнет или будет повреждена. Дублирование ДНК также является преимуществом. Деление частиц внутри организма позволяет быстрее расти и восстанавливаться поврежденным тканям.
Во время своего деления новая клетка копирует прежнюю, что позволяет сохранить благоприятные черты в следующих поколениях, а также со временем их усовершенствовать. Другими словами, дублирование позволяет сохранить и адаптировать черты, которые улучшат выживание или пригодность организма, особенно в царстве животных, подцарстве многоклеточных.

Недостатки многоклеточных
У сложных организмов имеются и недостатки. Например, они подвержены различным заболеваниям, возникающим из-за комплексного биологического состава и функций. У простейших, наоборот, не хватает развитых систем органов. Это означает, что риски опасных болезней у них сведены к минимуму.
Важно отметить, что в отличие от многоклеточных, примитивные особи обладают способностью к бесполому размножению. Это помогает им не тратить ресурсы и энергию на поиски партнера и сексуальную деятельность.
Простейшие организмы также обладают способностью принимать энергию путем диффузии или осмоса. Это освобождает их от необходимости передвижения для поиска пищи. Практически все может стать потенциальным источником пищи для одноклеточного существа.
Позвоночные и беспозвоночные
Всех без исключения входящих в подцарство многоклеточных существ классификация делит на два вида: позвоночных (хордовых) и беспозвоночных.
У беспозвоночных нет твердого каркаса, в то время как хордовые имеют хорошо развитый внутренний скелет хряща, кости и высокоразвитый мозг, который защищен черепом. Позвоночные имеют прекрасно развитые органы чувств, дыхательную систему с жабрами или легкими и развитую нервную систему, что еще больше отличает их от более примитивных собратьев.
Оба типа животных живут в различных местах обитания, но хордовые, благодаря развитой нервной системе, могут адаптироваться к суше, морю и воздуху. Тем не менее, беспозвоночные также встречаются в широком диапазоне, от лесов и пустынь до пещер и грязи морского дна.
На сегодняшний день выявлено почти два миллиона видов подцарства многоклеточных беспозвоночных животных. Эти два миллиона составляют около 98 % от всех живых существ, то есть 98 из 100 видов проживающих в мире организмов — беспозвоночные. Человеческие особи относятся к семейству хордовых.
Позвоночные подразделяются на рыб, земноводных, рептилий, птиц и млекопитающих. Не имеющие позвоночника животные представляют такие типы, как членистоногие, иглокожие, черви, кишечнополостные и моллюски.
Одним из самых главных различий между этими видами является их размер. Беспозвоночные, такие как насекомые или кишечнополостные, малы и медлительны, потому что не могут развить крупное тело и сильные мышцы. Есть несколько исключений, таких как кальмар, который может достигать 15 метров в длину. Позвоночные имеют универсальную систему поддержки, а потому могут быстрее развиваться и становиться крупнее, чем беспозвоночные.
Хордовые имеют также высокоразвитую нервную систему. С помощью специализированной связи между нервными волокнами, они могут реагировать очень быстро на изменения в окружающей среде, что дает им несомненное преимущество.
По сравнению с позвоночными, большинство животных, не имеющих хребта, используют простую нервную систему и ведут себя почти полностью инстинктивно. Подобная система работает хорошо большую часть времени, хотя эти существа часто неспособны учиться на своих ошибках. Исключениями являются осьминоги и их близкие родственники, которые считаются одними из самых умных животных в мире беспозвоночных.
У всех хордовых, как нам известно, имеется позвоночник. Однако особенностью у подцарства многоклеточных беспозвоночных животных является сходство с их сородичами. Оно заключается в том, что на определенном этапе жизни позвоночные также имеют гибкий опорный стержень, нотохорд, который впоследствии становится позвоночником. Первая жизнь развивалась в виде одиночных клеток в воде. Беспозвоночные были начальным звеном эволюции других организмов. Их постепенные изменения привели к появлению сложных существ с хорошо развитым скелетом.

Кишечнополостные животные
Сегодня насчитывается около одиннадцати тысяч видов кишечнополостных. Это одни из самых древнейших сложных животных, появившихся на земле. Самых маленьких из кишечнополостных невозможно увидеть без микроскопа, а самая большая известная медуза — 2,5 метра в диаметре.
Итак, давайте подробнее познакомимся с подцарством многоклеточных, типом кишечнополостные. Описание основных характеристик мест обитания можно определить наличием водной или морской среды. Они живут одиночно или в колониях, которые могут свободно передвигаться или жить на одном месте.
Форма тела кишечнополостных называется «мешком». Рот соединяется со слепым мешком, который называется «гастроваскулярной полостью». Этот мешок функционирует в процессе пищеварения, газообмена и действует как гидростатический скелет. Единственное отверстие служит как ртом, так и задним проходом. Щупальца — длинные, полые структуры, используются для перемещения и захвата пищи. Все кишечнополостные имеют щупальца, покрытые присосками. Они оснащены специальными клетками — немоцистами, которые могут впрыскивать токсины в свою жертву. Присоски также позволяют захватывать крупную добычу, которую животные помещают в рот путем втягивания щупалец. Нематоцисты отвечают за ожоги, которые некоторые медузы наносят людям.
Животные подцарства многоклеточные, типа кишечнополостные обладают как внутриклеточным, так и внеклеточным пищеварением. Дыхание происходит путем простой диффузии. У них имеется сеть нервов, которые распространяются по всему телу.
Многие формы проявляют полиморфизм, то есть разнообразие генов, в котором различные типы существ присутствуют в колонии для различных функций. Эти особи называются зооидами. Воспроизводство можно называть беспорядочным (внешнее почкование) или половым (формирование гамет).
Медузы, например, производят яйцеклетки и сперматозоиды, а затем выпускают их в воду. Когда яйцо оплодотворено, оно развивается в свободно плавающую личинку с ресничками, называемую «планлой».
Типичными примерами подцарства Многоклеточные типа кишечнополостные являются гидры, обелия, португальский кораблик, парусница, медуза-аурелия, медуза-кочан, актинии, кораллы, морское перо, горгонарии и т. д.

Растения
В подцарстве Многоклеточные растения - это эукариотические организмы, способные питаться в процессе фотосинтеза. Водоросли изначально считались растениями, но теперь они относятся к протистам — особой группе, которая исключена из всех известных видов. Современное определение растений относится к организмам, которые живут в основном на суше (а иногда и в воде).
Другой отличительной особенностью растений является зеленый пигмент — хлорофилл. Он используется для поглощения солнечной энергии в процессе фотосинтеза.
У каждого растения есть гаплоидные и диплоидные фазы, которые характеризуют его жизненный цикл. Он называется чередованием поколений, потому что все фазы в нем являются многоклеточными.
Вероятно, 700-900 млн лет назад на Земле появились первые многоклеточные животные и растения. У растений возникновение многоклеточного уровня организации, вероятно, произошло на основе дифференциации лентообразных колоний, образовавшихся путем бокового срастания прикрепленных нитчатых форм или благодаря делению клеток последних в двух взаимно перпендикулярных направлениях (в одной плоскости). У колоний, прикрепленных одним концом к субстрату, различные участки находились в разных условиях по отношению к падающему свету, субстрату и водной среде. В связи с этим естественный отбор благоприятствовал возникновению некоторой дифференциации частей колонии. Первым шагом было возникновение полярности колонии; на одном ее конце выделялись клетки, служившие для прикрепления к субстрату (для них характерно ослабление фотосинтеза, потеря способности к делению), на другом же конце - верхушечные клетки, интенсивно делившиеся и образовавшие своего рода «точку роста» колонии. Естественный отбор благоприятствовал приобретению клетками колонии способности делиться в разных направлениях; это приводило к ветвлению, что увеличивало поверхность колонии. Деление клеток вдоль трех взаимно перпендикулярных осей или переплетение отдельных нитей вело к возникновению многослойного «объемного» тела. В процессе его дальнейшей дифференциации сформировались многоклеточные органы, выполнявшие разные функции (фиксация на субстрате, фотосинтез, размножение). Одновременно между разными клетками растения складывалась определенная взаимозависимость, что, собственно говоря, и знаменует достижение многоклеточного уровня организации.
У животных активный образ жизни требовал более совершенной и сложной дифференциации организма, чем у растений. Сложность организации многоклеточных животных (Metazoa) и разнообразие ее конкретных форм стимулировали разработку различных гипотез о происхождении Metazoa .
Первая из них берет начало в работах Э. Геккеля, который в разработке своей теории гастреи основывался на сформулированном им биогенетическом законе, согласно которому онтогенез данного вида организмов представляет собой сжатое и сокращенное повторение (рекапитуляцию) хода филогенеза его предков (подробнее см. в ч. ГУ). В соответствии с этим Э. Геккель полагал, что филогенез древнейших Metazoa в определенной степени повторяется в онтогенезе современных низших многоклеточных животных (рис. 28). Согласно Геккелю, предками Metazoa были колониальные простейшие, обладавшие сферическими колониями с однослойной стенкой, подобными бластуле - одной из ранних
Рис. 28.
а - бластула; 6 - гаструляция; в-г - гаструла (внешний вид и продольный
разрез) стадий эмбрионального развития современных многоклеточных животных. Геккель назвал эту гипотетическую предковую форму «бластеей». При направленном плавании сферическая колония - бластея - ориентировалась одним полюсом вперед, как это наблюдается и у современных колониальных простейших, например у Volvox. Согласно Геккелю, на переднем полюсе колонии возникло впячивание ее стенки внутрь, подобно тому, как это происходит при инвагинационной гаструляции в онтогенезе некоторых современных Metazoa. В результате образовался многоклеточный организм - «гастрея», стенка тела которого состоит из двух слоев, экто- и энтодермы. Энтодерма окружает внутреннюю полость - первичный кишечник, открытый наружу единственным отверстием - первичным ртом. Организация гастреи соответствует принципиальному плану строения кишечнополостных (тип Coelenterata), которых Геккель и рассматривал как наиболее примитивных многоклеточных животных.
И. И. Мечников обратил внимание на то, что у примитивных кишечнополостных гаструляция происходит не путем инвагинации (впячивания одного полюса однослойного зародыша - бластулы), что характерно для более высокоорганизованных групп, а посредством миграции некоторых клеток из однослойной стенки тела внутрь (рис. 29). Там они образуют рыхлое скопление, позднее организующееся в виде стенок гастральной полости, которая прорывается наружу ротовым отверстием. Такой способ гаструляции гораздо проще, чем инвагинация, так как не требует сложного направленного и координированного смещения целого пласта клеток, и, вероятно, примитивнее инвагинации. В связи с этим Мечников модифицировал гипотезу Геккеля следующим образом. В сфероидной колонии простейших - жгутиконосцев клетки ее однослойной стенки, захватывавшие (фагоцитировав-

Рис. 29. Гаструляция зародыша гидроидного полипа Stomateca (из И.А. Иоффа) шие) пищу, мигрировали для ее переваривания внутрь, в полость колоний (подобно миграции клеток будущей энтодермы в процессе гаструляции кишечнополостных). Эти клетки образовали рыхлое внутреннее скопление - фагоцитобласт, функцией которого стало обеспечение всего организма пищей, включая ее переваривание и распределение, тогда как поверхностный слой клеток - кинобласт - осуществлял функции защиты и движения организма. Для захвата новых пищевых частиц клеткам фагоци- тобласта, по мысли Мечникова, не было необходимости возвращаться в поверхностный слой: располагаясь непосредственно под кинобластом, клетки фагоцитобласта захватывали пищевые частицы псевдоподиями, выдвигаемыми наружу в промежутках между клетками фагоцитобласта. Эта гипотетическая стадия эволюции Metazoa была названа Мечниковым фагоцителлой (или паренхимеллой); ее строение соответствует таковому паренхимулы, личинки некоторых кишечнополостных и губок. В дальнейшем как приспособление к повышению активности питания у потомков фагоцителлы произошла эпителизация фагоцитобласта с образованием первичного кишечника и возникновением ротового отверстия в том месте, где происходила преимущественная миграция клеток внутрь. По мнению некоторых ученых, это место, вероятно, соответствовало заднему по направлению движения полюсу тела, где при плавании возникают завихрения водяного потока, и поэтому условия наиболее благоприятны для захвата пищевых частиц. Гипотеза Мечникова, как и гипотеза Геккеля, рассматривает в качестве наиболее примитивных многоклеточных животных кишечнополостных и губок.
Важные сведения для понимания ранних этапов эволюции Metazoa были получены при изучении крайне примитивного многоклеточного животного трихоплакса (Trichoplax adhaerens), обнаруженного в Красном море Ф. Шульце еще в 1883 г., но детально исследованного лишь в 1970-е гг. нашего века К. Греллом и А. В. Ивановым. Трихоплакс (рис. 30) имеет уплощенное тело, лишенное полярности. Поверхность тела, обращенная вверх, покрыта плоским, а нижняя - цилиндрическим мерцательным эпителием. Внутри, между эпителиальными слоями, соответствующими кинобласту, находится полость с жидким содержимым, в котором располагаются веретеновидные и звездчатые клетки. Эти последние можно рассматривать как фагоцитобласт. Размножается трихоплакс бесполым способом - делением и почкованием. А. В. Иванов указал, что трихоплакс представляет собой как бы живую модель фагоцителлы, и предложил выделить эту форму в особый тип животных Phagocytellozoa. По-видимому, трихоплакс подкрепляет позиции гипотезы фагоцителлы И. И. Мечникова Однако по современным представлениям непосредственными

Рис. 30.
и - изменения формы тела одной особи (по Ф. Шульце); б - разрез, перпендикулярный краям тела (по А. В. Иванову): 1 - амебоидные клетки; 2 - спинной эпителий; 3 - веретеновидные клетки; 4 - жировые включения; 5 - пищеварительные вакуоли; 6 - брюшной эпителий
потомками фагоцителлозой среди многоклеточных были не кишечнополостные, а примитивные червеобразные животные, близкие по уровню организации к плоским ресничным червям - тур- белляриям.
Первые ископаемые следы жизнедеятельности червеобразных многоклеточных животных известны из позднерифейских отложений. В вендское время (650-570 млн лет назад) существовали уже разнообразные животные, вероятно принадлежавшие к различным типам. Немногочисленные отпечатки мягкотелых вендских животных известны из разных районов всех континентов земного шара, кроме еще малоисследованной Антарктиды. Ряд интересных находок был сделан в позднепротерозойских отложениях на территории России - на Кольском полуострове, в Архангельской области, на реке Мая и на Оленекском поднятии в Якутии и т. д.

Рис. 31.
1-10 - кишечнополостные (/ - Ediacara; 2 - Beltanella; 3 - Mcdusinitcs; 4 - Mawsonites; 5-6- Cyclomedusa; 7 - Conomedusites; 8 - Rangea; 9- Arborea; 10 - Pteridinium); 11-14 - плоские и кольчатые черви (11 - Spriggina; 12-14 - Dickinsonia); 15-16 - членистоногие (15 - Parvancorina; 16 - Praecambridium); 17 - иглокожее Tribrachidium; 18 - шарообразные студенистые организмы
Наиболее известна богатая позднепротерозойская фауна, обнаруженная в Центральной Австралии в районе Эдиакары к северу от г. Аделаида. Исследовавший эту фауну М. Глесснер считает, что она включает несколько десятков видов очень разнообразных многоклеточных животных, относящихся к разным типам (рис. 31). Большинство форм принадлежат, вероятно, к кишечнополостным. Это медузоподобные организмы, вероятно «парившие» в толще воды (Ediacara flindersi, Beltanella gilesi, Medusinites asteroides и др.), и прикрепленные к морскому дну полипоидные формы, одиночные или колониальные, напоминающие современных кораллов альционарий, или морские перья (Rangea longa, Arborea arborea, Pteridinium simplex и др.). Замечательно, что все они, как и другие животные эдиакарской фауны, лишены твердого скелета.
Кроме кишечнополостных в составе эдиакарской фауны найдены остатки червеобразных животных, причисляемых к плоским и кольчатым червям (Spriggina floundcri и разные виды Dickinsonia). Некоторые виды организмов интерпретируют как возможных предков членистоногих (Praecambridium sigillum, напоминающий по характеру сегментации тела трилобитов и хелицеровых) и иглокожих (Tribrachidium heraldicum с телом дисковидной формы, на плоской поверхности которого выступают три валика, и Arkaria
adami с пятилучевой звездообразной впадиной на ротовой стороне тела и с подобием амбулакральных желобков). Наконец, имеется целый ряд ископаемых организмов неизвестной таксономической принадлежности.
Многие вендские организмы были обнаружены также в вендских отложениях разных районов России: медузоподобные эдиа- кария и медузинитес - на полуострове Рыбачьем, птеридиниум - на севере Якутии, напоминающая сприггину вендия - в районе Яренска Архангельской области и т. д. Местонахождения вендской фауны, по богатству не уступающие эдиакарскому, были найдены на реке Сюзьма на Онежском полуострове и на Зимнем берегу Белого моря. Здесь были обнаружены ископаемые остатки свыше 30 видов бесскелетных многоклеточных животных, размеры которых варьировали от 3 мм до 30 см. Среди них вероятные представители кишечнополостных, плоских и кольчатых червей, членистоногих, иглокожих, а также ряд форм, принадлежащих к каким-то неизвестным группам. Вообще родство вендских организмов с современными группами, большинство из которых достоверно известно начиная с кембрия, остается проблематичным - различия очень велики, и некоторые исследователи считают, что известные ныне вендские организмы не связаны прямым родством с более поздними кембрийскими, а представляют слепые эволюционные ветви.
М. А. Федонкин, исследовавший Беломорскую фауну вендских животных, полагает, что некоторые из этих организмов обладают признаками нескольких разных типов животных и могут представлять исходные формы, занимающие промежуточное положение. Федонкин обратил также внимание на сходство ряда вендских организмов с личиночными стадиями некоторых современных животных, хотя вендские организмы имеют значительно более крупные размеры, чем соответствующие личинки. При всем разнообразии планов строения тела у организмов вендской фауны («вендобионтов») их объединяют некоторые общие черты организации: отсутствие скелета, конечностей, вероятно, также дыхательных и пищеварительных органов. Многие вендобионты вели неподвижный прикрепленный образ жизни. Некоторые исследователи полагают, что вендские организмы питались осмотически через поверхность тела или же с помощью живших в их теле фото- или хемосинтезирующих симбионтов - одноклеточных водорослей и бактерий.
Хотя среди вендских животных преобладают мягкотелые бес- скелетные формы, вероятно, в те времена существовали уже и немногие виды, обладавшие раковиной. Такова, например, Cloudina, имевшая простую трубчатую раковину, состоявшую из органического вещества и кальцита. Клаудина была обнаружена в карбонатных породах, которые переслаиваются с отложениями, содержащими остатки эдиакарской фауны мягкотелых животных.
Все эти данные указывают на широкое распространение фаун мягкотелых животных в вендское время. Накопление материалов по вендским ископаемым организмам позволило некоторым исследователям поставить вопрос о расширении рамок фанерозоя, с включением в его состав «эдиакария» - периода, охватывающего промежуток времени от 670 до 550 млн лет назад (в приведенной на с. 149 схеме геохронологии этот период соответствует венду в составе протерозоя).
Поскольку вендская фауна столь разнообразна и включает довольно высокоорганизованных животных, очевидно, что до ее возникновения эволюция Metazoa продолжалась уже очень долго. Вероятно, многоклеточные животные появились значительно раньше - где-то в промежутке 700-900 млн лет назад .
Таким образом, в позднем протерозое (600-650 млн лет назад) уже существовали такие группы многоклеточных животных, как губки, кишечнополостные, плоские и кольчатые черви и даже, возможно, предки членистоногих. Судя по достигнутому уровню организации, можно предполагать, что к этому времени обособились также эволюционные стволы нитчатых червей (тип Nemathelminthes), предков моллюсков и предков вторичноротых животных - олигомерных червей.
Докембрийский филогенез Metazoa можно гипотетически представить следующим образом (рис. 32). От колониальных жгутиковых (по мнению ряда авторов - от гетеротрофных форм, принадлежавших к отряду Protomonadida) путем дифференциации и интеграции колонии, с миграцией внутрь колонии клеток фа- гоцитобласта на заднем полюсе тела, возникли первые многоклеточные животные, организация которых соответствовала фа- гоцителле (по И. И. Мечникову). Мало изменившимися потомками этих древнейших многоклеточных являются современные Phagocytellozoa (Trichoplax adhaerens). Примитивные многоклеточные были свободноплавающими (за счет работы мерцательного эпителия - кинобласта) животными, питавшимися различными микроорганизмами - простейшими и одноклеточными водорослями.
При дальнейшем развитии приспособлений к активному питанию происходила постепенная эпитслизация фагоцитобласта, т. е.

Рис. 32.
преобразование рыхлого скопления клеток в организованный клеточный пласт - эпителий кишечника. Эпителизация фагоцито- бласта, вероятно, началась с развития на заднем по движению полюсе тела постоянного ротового отверстия. Как отметил К. В. Беклемишев, на этой стадии филогенеза организм стал питаться как целое, а не как совокупность отдельных самостоятельно фагоцитирующих клеток. Вероятно, к этому времени сформировалась и интегрирующая организм нервная система в виде эпителиального нервного сплетения. Активное плавание требовало способности ориентироваться в пространстве и координировать работу всех органов. Для осуществления этих функций на аборальном (противоположном ротовому отверстию) полюсе тела возник нейро-рецепторный комплекс, включавший нервный ганглий, осязательные щетинки и статоцист (орган равновесия). Подобный аборальный орган имеется у современных гребневиков (тип Ctenophora), а также у свободноплавающих личинок очень многих групп животных: плоских и кольчатых червей, моллюсков, членистоногих, полухордовых, иглокожих и др. Эту гипотетическую стадию филогенеза древних Metazoa можно назвать «стомо- фагоцителлой» (подчеркивая эпителизацию лишь ротового отдела фагоцитобласта).
Возможно, на этой стадии филогенеза произошла первая крупная дивергенция филогенетического ствола древних многоклеточных, связанная с тем, что некоторые группы этих животных перешли к освоению морского дна, другие же продолжали совершенствовать приспособления к активной жизни в толще воды.
Современные низшие плоские черви - бескишечные турбел- лярии (Acoela) в целом сохранили тот уровень организации, который, вероятно, был характерен для древнейших многоклеточных, впервые перешедших к освоению подвижного образа жизни на дне водоемов. От вендских представителей этих турбеллярий могли возникнуть филогенетические стволы, ведущие к другим группам плоских червей, к нитчатым червям и к предкам кольчатых червей (протоаннелидам). От протоаннелид обособились, с одной стороны, предки моллюсков, с другой - предки членистоногих. У всех этих групп произошла дальнейшая дифференциация фагоцитобласта. У низших червей эпителизировалась лишь его центральная часть, что привело у плоских червей к формированию разветвленного кишечника с единым отверстием - «ртом», ведущим во внешнюю среду, а у нитчатых червей - к образованию сквозного кишечника с ротовым и анальным отверстиями. У высших групп (кольчатые черви, моллюски и членистоногие) эпитслизировался весь фагоцитобласт: не только его центральная часть (энтодермальный кишечник), но и периферическая (мезодерма и ее производные). Последнее привело к развитию вторичной полости тела - целома, стенки которой образованы мезодер- мальным целомическим эпителием. Более примитивные представители кольчатых червей, моллюсков и членистоногих обладают характерной личиночной стадией - трохофорой. В связи с этим указанные группы иногда объединяют под названием Trochozoa.
У тех потомков стомофагоцителлы, которые продолжали совершенствовать адаптации к жизни в толще воды, также произошла эпителизация центрального и отчасти периферического фагоцитобласта: возникла гастральная полость (первичный кишечник) и ее периферические ветви (гастроваскулярные каналы). К этому уровню организации среди современных животных ближе всего стоят гребневики, вероятно сохранившие примитивный образ жизни в толще воды. От их позднепротерозойских предков, которых можно назвать «проктенофорами», с переходом к прикрепленной жизни на морском дне возникли стрекающие кишечнополостные (тип Coelenterata, или Cnidaria).
Другие филогенетические линии, ответвившиеся от проктено- фор, также осваивали морское дно, но с развитием приспособлений к активному передвижению по субстрату, подобно тур- белляриям и их потомкам, но на другом исходном уровне организации. У этих форм в результате завершения эпителизации периферического фагоцитобласта также образовалась вторичная полость тела - целом, но возникла она совершенно другим способом, чем у Trochozoa. В онтогенезе животных, происходящих от проктенофор, вторичная полость тела обособляется от первичного кишечника, как его боковые карманообразные выпячивания (первоначально было, вероятно, три пары таких выпячиваний), которые затем отшнуровываются от стенок кишки (рис. 33). Такой способ развития целома получил название энтероцельного - в отличие от схи- зоцельного способа, характерного для Trochozoa, у которых целом возникает в результате появления полостей внутри скоплений мезодермальных клеток, без всякой связи с первичным кишечником. О. и Р. Гертви- ги и И. И. Мечников обосновали гипотезу, согласно которой энтероцельный целом возник в эволюции из гастроваскулярных каналов проктенофорных предков (энтероцельная теория происхождения целома). Энтероцельный целом характерен для типов погонофор (Pogonophora), щетинкочелюстных (Chaeto- gnatha), плеченогих (Brachiopoda), мшанок (Bryozoa) и ряда других, в том числе группы так называемых вторичноротых животных (Deuterostomia), объединяющей типы хордовых (Chordata), иглокожих (Echinodermata) и полухордовых (Hemichordata). У вторичноротых животных имеется много общего, в частности особое положение дефинитивного (присущего взрослым организмам) рта, возникающего на полюсе тела, противоположном первичному эмбриональному рту - бла- стопору. На месте же последнего развивается анальное отверстие. Вторичноротые, несомненно, имеют общее происхождение; в качестве их предков указывают гипотетическую группу олигомерных червей, тело которых было разделено на три отдела, имелся вторичный рот и энтероцельный целом. Среди современных вторичноротых к уровню организации олигомерных червей ближе всего стоят свободноживущие полухордовые, представителем которых является желудевый червь (Balanoglossus).
Рис. 33.
I - эктодерма; 2 - энтодерма; 3 - мезодерма; 4 - первичный кишечник; 5 - целомические карманы; 6 - нервная пластинка; 7- целом; 8 - вторичный кишечник; 9 - нервная трубка; 10 - хорда
Особое положение среди многоклеточных животных занимают губки (тип Porifera, или Spongia). Эта группа характеризуется очень примитивным общим уровнем организации: губки, по существу, не имеют эпителизованного фагоцитобласта, упорядоченного внутреннего строения, настоящего кишечника, нервной системы, рецепторов и т. д. Губки отличаются от всех остальных Metazoa чрезвычайно своеобразным онтогенезом, в ходе которого происходит инверсия зародышевых листков (экто- и энтодерма, так сказать, меняются местами). Новейшие данные молекулярных исследований показали, что губки имеют общее происхождение со всеми многоклеточными. Вероятно, они представляют собой очень раннюю боковую ветвь, обособившуюся на уровне фагопителлы. Древнейшие ископаемые остатки губок известны из вендских (эдиа- карских) отложений в Австралии.
- См.: Иванов А.В. Происхождение многоклеточных. - Л., 1968.
- Недавно появились первые данные об ископаемых остатках многоклеточных животных, найденных в Канаде и в Китае в породах, имеющих такой геологический возраст и отделенных от пород с остатками эдиакарских организмовслоями ледниковых отложений (тиллитов).
Живой мир наполнен головокружительным множеством живых существ. Большинство организмов состоят только из одной клетки и не видимы невооруженным глазом. Многие из них становятся заметными исключительно под микроскопом. Другие, такие как кролик, слон или сосна, а также человек, сделаны из многих клеток, и эти многоклеточные организмы также в огромном количестве населяют весь наш мир.
Строительные блоки жизни
Структурными и функциональными единицами всех живых организмов являются клетки. Их еще называют строительными блоками жизни. Все живые организмы состоят из клеток. Эти структурные единицы были открыты Робертом Гуком еще в 1665 году. В организме человека насчитывается около ста триллионов клеток. Размер одной составляет около десяти микрометров. Ячейка содержит клеточные органеллы, которые контролируют ее активность.
Существуют одноклеточные и многоклеточные организмы. Первые состоят из одной клетки, например бактерии, а вторые включают растения и животных. Количество ячеек зависит от вида. Размер большинства клеток растений и животных клетках составляет от одного до ста микрометров, поэтому они видны под микроскопом.

Одноклеточные организмы
Эти крошечные существа состоят из одной клетки. Амебы и инфузории являются самыми старыми формами жизни, которые существовали еще около 3,8 миллиона лет назад. Бактерии, археи, простейшие, некоторые водоросли и грибы являются основными группами одноклеточных организмов. Существует две основные категории: прокариоты и эукариоты. Они также различаются по размеру.
Самые маленькие составляют около трехсот нанометров, а некоторые могут достигать размеров до двадцати сантиметров. Такие организмы обычно имеют реснички и жгутики, которые помогают им при перемещении. Они имеют простой корпус с базовыми функциями. Размножение может быть как бесполое, так и половое. Питание осуществляется обычно в процессе фагоцитоза, где частицы еды поглощаются и хранятся в специальных вакуолях, которые присутствуют в организме.

Многоклеточные организмы
Живые существа, состоящие из более чем одной клетки, называются многоклеточными. Они состоят из единиц, которые идентифицируются и присоединяются друг к другу, образуя сложные многоклеточные организмы. Большинство из них видны невооруженным глазом. Такие организмы, как растения, некоторые животные и водоросли, появляются из одной клетки и вырастают в многоцепочечные организации. Обе категории живых существ, прокариоты и эукариоты, могут проявлять многоклеточность.

Механизмы возникновения многоклеточности
Существует три теории для обсуждения механизмов, с помощью которых может возникнуть многоклеточность:
- Симбиотическая теория утверждает, что первая клетка многоклеточного организма возникла из-за симбиоза различных видов одноклеточных, каждый из которых выполняет различные функции.
- Синцитиальная теория утверждает, что многоклеточный организм не смог бы развиться из одноклеточных существ с несколькими ядрами. Такие простейшие, как инфузория и слизистые грибы, имеют несколько ядер, тем самым поддерживая эту теорию.
- Колониальная теория утверждает, что симбиоз многих организмов одного и того же вида приводит к эволюции многоклеточного организма. Она была предложена Геккелем в 1874 году. Большинство многоклеточных образований происходит вследствие того, что клетки не могут отделиться после процесса деления. Примерами, подтверждающими эту теорию, являются водоросли вольвокс и эудорина.

Преимущества многоклеточности
Какие организмы - многоклеточные или одноклеточные - имеют больше преимуществ? На этот вопрос ответить достаточно сложно. Многоклеточность организма позволяет ему превышать предельные размеры, увеличивает сложность организма, позволяя дифференцировать многочисленные клеточные линии. Размножение происходит преимущественно половым путем. Анатомия многоклеточных организмов и процессы, которые в них происходят, являются достаточно сложными из-за наличия различных типов клеток, контролирующих их жизнедеятельность. Возьмем, к примеру, деление. Этот процесс должен быть точным и слаженным, чтобы предотвратить ненормальный рост и развитие многоклеточного организма.

Примеры многоклеточных организмов
Как уже говорилось выше, многоклеточные организмы бывают двух видов: прокариоты и эукариоты. К первому относят в основном бактерий. Некоторые цианобактерии, такие как чара или спирогира, являются также многоклеточными прокариотами, иногда их называют еще колониальными. Большинство эукариотических организмов также состоят из множества единиц. Они имеют хорошо развитую структуру тела, и у них есть специальные органы для выполнения определенных функций. Большинство хорошо развитых растений и животных являются многоклеточными. Примерами могут быть практически всех виды голосеменных и покрытосеменных растений. Почти все животные являются многоклечточными эукариотами.

Особенности и признаки многоклеточных организмов
Существует масса признаков, по которым можно с легкостью определить, является ли организм многоклеточным или нет. Среди можно выделить следующие:
- У них достаточно сложная организация тела.
- Специализированные функции выполняют различные клетки, ткани, органы или системы органов.
- Разделение труда в организме может быть на клеточном уровне, на уровне тканей, органов и уровне систем органов.
- В основном это эукариоты.
- Травмы или гибель некоторых клеток глобально не влияет на организм: пораженные клетки будут заменены.
- Благодаря многоклеточности организм может достигать больших размеров.
- По сравнению с одноклеточными у них большая продолжительность жизненного цикла.
- Основной тип размножения - половой.
- Дифференциация клеток свойственна только многоклеточным.
Как растут многоклеточные организмы?
Все существа, от маленьких растений и насекомых до больших слонов, жирафов и даже людей, начинают свой путь как единичные простые клетки, называемые оплодотворенными яйцами. Чтобы вырасти в большой взрослый организм, они проходят через несколько определенных этапов развития. После оплодотворения яйца начинается процесс многоклеточного развития. На протяжении всего пути происходит рост и многократное деление отдельных ячеек. Эта репликация в конечном итоге создает конечный продукт, который является сложным, полностью сформированным живым существом.
Разделение клеток создает ряд сложных моделей, определяющихся геномами, которые являются практически идентичными во всех клетках. Это разнообразие приводит к экспрессии генов, которая контролирует четыре стадии развития клеток и эмбрионов: пролиферацию, специализацию, взаимодействие и движение. Первая включает в себя репликацию многих клеток из одного источника, вторая имеет отношение к созданию клеток с выделенными, определенными характеристиками, третья включает в себя распространение информации между ячейками, а четвертая отвечает за размещение клеток по всему телу для образования органов, тканей, костей и других физических характеристик развитых организмов.
Несколько слов о классификации
Среди многоклеточных существ выделяют две большие группы:
- беспозвоночные (губки, кольчатые черви, членистоногие, моллюски и другие);
- хордовые (все животные, у которых есть осевой скелет).
Важным этапом за всю историю планеты стало появление многоклеточности в процессе эволюционного развития. Это послужило мощным толчком для увеличения биологического разнообразия и его дальнейшего развития. Главным признаком многоклеточного организма является четкое распределение клеточных функций, обязанностей, а также установка и налаживание устойчивых и прочных контактов между ними. Другими словами, это многочисленная колония клеток, которая в силах сохранять фиксированное положение на протяжении всего жизненного цикла живого существа.